

5.3.2. Морфофункціональна характеристика нейронів
Нейрони складаються із тіла (соми), одного або кількох відростків (дендритів), по яких нервовий імпульс надходить до тіла нейрона, й одного відростка (аксона), по ньому нервовий імпульс поширюється від одного нейрона до іншого або до відповідних органів з утворенням нервових
волокон. Тіла нейронів розташовані у сірій речовині мозку і виконують трофічну функцію. Унікальність нейронів полягає у тому, що незабаром після народження людини вони втрачають здатність до фізіологічної регенерації шляхом поділу. Самовідновлення їх відбувається лише на рівні субклітинних структур або окремих молекул.
Діаметр аксонів залежить від величини нейрона (від 5 до 100 мкм): у малих нейронів - близько 1 мкм, у великих - до б мкм. Місце нейрона, звідки відходить аксон, називається аксонним горбком.
Тіло нейрона покрите типовою плазматичною мембраною. На ній наявні всі види структур, що забезпечують трансмембранний транспорт і підтримання градієнтів іонних концентрацій.
Градієнти концентрації іонів, що створюються завдяки діяльності відповідних помп між внутрішнім вмістом клітини і міжклітинною рідиною, забезпечують існування потенціалів спокою та виникнення потенціалу дії. Більшість функціональних проявів нейрона зумовлені змінами потенціалів мембрани. МП спокою не у всіх нейронах однаковий і коливається від -90 до -40-60 мв. У великих нейронах він значно вищий, ніж у дрібних.
Мембрана аксонного горбка дещо відрізняється від інших відділів соми нейрона. Тут мембранний потенціал спокою менший (близько -60 мв), тобто це найзбудливіша частина нейрона. Саме в цьому місці уперше виникає сумарний потенціал дії.
Класифікація нейронів. Розрізняють аферентні (рецепторні, чутливі), проміжні (вставні, контактні) та еферентні (рухові) нейрони. Аферентні нейрони проводять чутливі імпульси від рецепторів до ЦНС. Це біполярні клітини, що мають два довгих відростки - один проводить імпульси від рецепторів до тіла, інший - від тіла до ЦНС. Тіла біполярних клітин розташовуються поза ЦНС (у спинномозкових гангліях або чутливих гангліях черепних нервів). Еферентні нейрони проводять імпульси до органів і тканин, їхні тіла (крім автономної нервової системи) розташовані в межах ЦНС. Проміжні нейрони найбільш численні. Вони передають нервові імпульси від аферентних до еферентних нейронів і один до одного, зв'язуючи різні нервові клітини між собою з утворенням нейронних сіток.
За своєю функцією проміжні нейрони можуть бути збудливими (вони активують інші нейрони, з якими контактують) і гальмівними (за їх активації відбувається зниження активності інших нейронів).
5.3.3. Синапси центральної нервової системи
Нервові клітини за допомогою своїх відростків функціонують у тісній взаємодії одна з одною. Передача нервових імпульсів від одного нейрона до інших здійснюється за допомогою синапсів. У результаті кожен нейрон контактує прямо або (найчастіше) опосередковано із сотнями, тисячами інших з утворенням нейронних сіток. Відстань між окремими синапсами приблизно однакова, тому їхня кількість на тілі нейрона насамперед ви-
значається розмірами соми: на малих клітинах їх до 5000, а на великих -до 200 000. Однак наявні й функціональні відмінності в кількості синапсів на тілі клітини: у чутливих нейронів синапсів менше, у вставних та еферентних - більше.
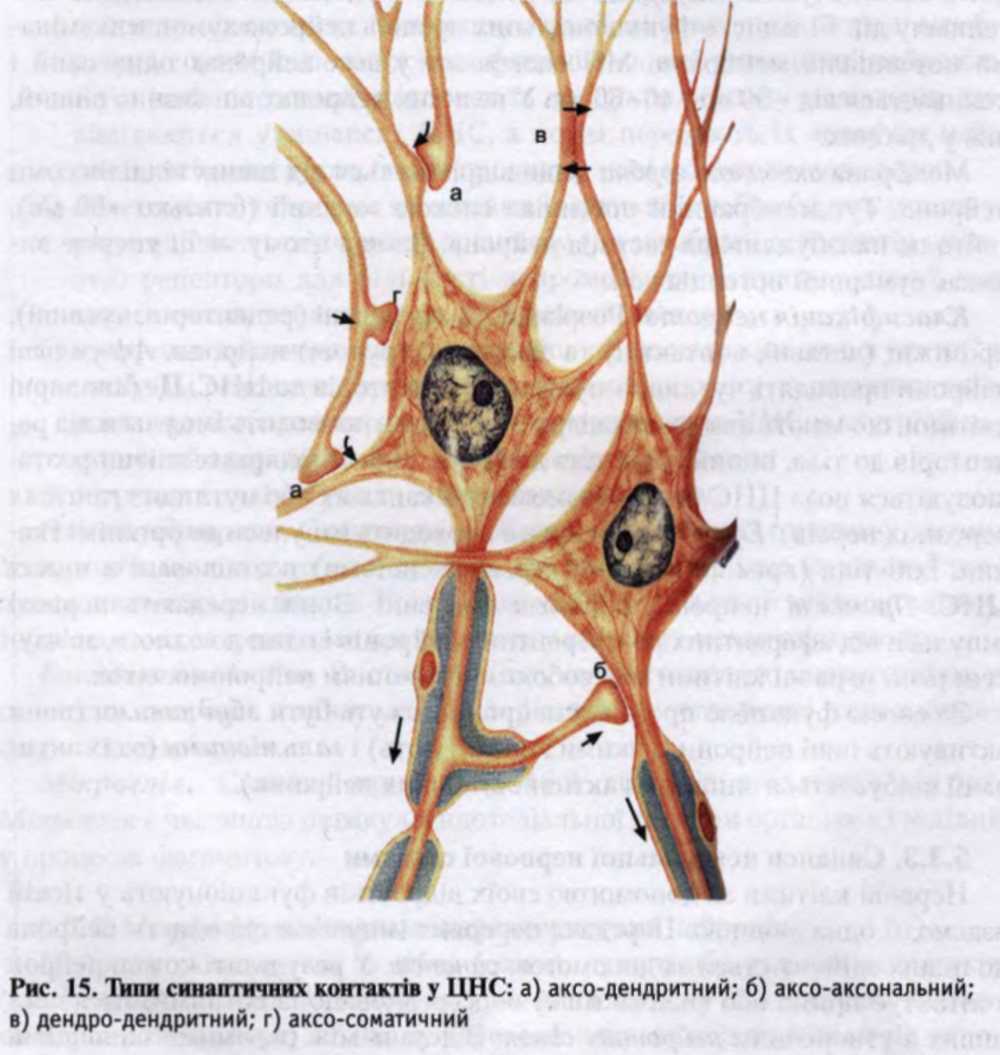
Класифікація синапсів. Залежно від розташування міжнейронних контактів розрізняють аксо-соматичні, аксо-дендритні, аксо-аксонні, денд-ро-дендритні, дендро-соматичні (рис. 15). За функцією синапси бувають збудливі, коли нервовий імпульс, що прийшов до нього, збуджує постси-наптичний нейрон, і гальмівні, коли такий імпульс призводить до пригнічення активності постсинаптичного нейрона. За механізмом передачі сигналу між нейронами синапси можуть бути хімічні та електричні.
Будова хімічного синапса
Переважна більшість синапсів ЦНС - хімічні. Вони збудовані приблизно так само, як і нервово-м'язові (рис. 12). їх будову можна розглянути на прикладі аксо-соматичного синапса. Аксон пресинаптичного нейрона підходить до тіла іншого (постсинаптичного) й утворює пресинаптичне закінчення (кінцеву бляшку). Концентрація іонів кальцію у пресинаптич-ному закінченні менша, ніж у навколишньому міжклітинному середовищі. Та частина мембрани пресинаптичного закінчення, що прилягає до тіла постсинаптичного нейрона, називається пресинаптичною мембраною. Частина мембрани тіла постсинаптичного нейрона у ділянці пресинаптичної бляшки називається постсинаптичною мембраною. Вона містить хемочут-ливі (лігандзалежні) натрієві канали, що відкриваються у разі дії певних хімічних речовин (лігандів). Між цими двома мембранами розміщена си-наптична щілина. У пресинаптичному закінченні у пухирцях діаметром близько 50 нм містяться біологічно активні сполуки, медіатори або ней-ротрансмітери. За допомогою медіаторів нервові імпульси передаються через синапс від одного нейрона на інший.
На постсинаптичній мембрані локалізовані хеморецептори, з якими може зв'язуватися медіатор, споріднений з ними, а це призводить до відкривання іонних каналів мембрани.
Нейротрансмітери
У ЦНС функцію трансмітерів виконує велика кількість біологічно активних речовин (близько ЗО). Належність синапсів до збудливих чи гальмівних визначається, з одного боку, специфікою медіаторів, аз іншого -видом рецепторів, які розташовані на постсинаптичній мембрані. Звичайно один і той же медіатор має на ній декілька рецепторів. Тому під час взаємодії одного і того ж трансмітера з різними рецепторами може виникати різний ефект - активація або гальмування постсинаптичного нейрона. Характер рецептора можна визначити не тільки за відмінністю ефекту, але й шляхом застосування хімічно активних речовин, що блокують передачу нервового імпульса через синапс (результат зв'язування з рецептором) або посилюють вплив трансмітера. Ці речовини можуть бути як ендогенного походження (утворюватися у самій ЦНС чи в інших органах і надходити до ЦНС через кров та лімфу), так і екзогенного (надходити із зовнішнього середовища) -сполуки, що надходять з їжею, лікарські препарати, різні отрути, токсини.
Трансмітери ЦНС залежно від хімічної будови можна розділити на чотири основні групи:
1. Аміни (ацетилхолін, норадреналін, адреналін, дофамін, серотонін).
2. Амінокислоти (гліцин, глутамін, аспарагінова кислота, гамааміно- масляна кислота (ГАМК) тощо).
3. Пуринові нуклеотиди.
4. Нейропептиди (гіпоталамічні ліберини і статини, оніоїдні пептиди,
вазопресин, речовина Р, холецистокінін, гастрин тощо). У деяких синапсах виявлені також простагландини. У багатьох нейронах може міститися не один, а два чи більше трансмітерів.
За своїм механізмом дії трансмітери можна поділити на два типи: іоно-тропні і метаботропні. Іонотропні трансмітери після взаємодії з рецепторами постсинаптичної мембрани змінюють проникність іонних каналів. На відміну від них метаботропні медіатори постсинаптичний вплив реалізують шляхом активації специфічних ферментів. У результаті в мембрані, а найчастіше - в цитозолі клітини активуються вторинні посередники (месенджери), які, у свою чергу, запускають каскади ферментативних реакцій у клітині або мембрані. Ці метаболічні зміни триваліші та глибші, ніж у разі дії іонотропних медіаторів. Вони можуть торкатися геному клітини, навіть передаватися спадково.
Метаботропною активністю володіють більшість нейропептидів та низка інших трансмітерів, наприклад аміни. У разі виділення разом з основним медіатором вони здатні модулювати (підсилювати чи послаблювати) його ефект або регулювати його вихід.
У результаті складних взаємодій, що відбуваються у ЦНС між трансмітерами і рецепторами, різними трансмітерами, ефект дії медіаторів може проявлятися по-різному. Так, ацетилхолін в одному випадку може викликати збудження, а в іншому - гальмування. Аналогічні ефекти характерні і для амінів. Серед амінокислот можна виділити ГАМК, що реалізує як пре-, так і постсинаптичне гальмування, гліцин, який реалізує постси-наптичне гальмування, та глутамінову кислоту, що передає збудження.
Механізм функціонування збуджувальних хімічних синапсів
Під впливом потенціалу дії відбувається деполяризація пресинаптич-ної мембрани, що зумовлює відкривання розташованих тут кальцієвих каналів. Іони кальцію пасивно (за градієнтом концентрації) надходять у пре-синаптичне закінчення, що сприяє наближенню міхурців з трансмітером до мембрани і, як наслідок, виділення його у синаптичну щілину.
Трансмітер, дифундуючи через синаптичну щілину, досягає постсинаптичної мембрани, де з'єднується з відповідним рецептором, у результаті чого відкриваються хемозбудливі канали і підвищується проникність мембрани для іонів або Са2+. Це зумовлює деполяризацію мембрани -виникнення внутрішнього струму, що не досягає критичного рівня деполяризації. Такий потенціал називається збуджувальним постсинаптинним потенціалом (ЗПСП). Величина утвореного внаслідок цього струму є настільки малою, що він не здатний поширюватися, доки подібні впливи не деполяризують усю мембрану. ЗПСП, зумовлені активністю в основному синаптичному контакті, невеликі, проте деполяризації, що виникають у
кожному з активних синапсів, додаються. Генерація ПД відбувається у результаті сумації ЗПСП. Цьому сприяють характеристики ЗПСП: порівняно велика його тривалість (наростання деполяризації 1-2 мс і спад - 10-12 мс), здатність поширюватися на сусідні ділянки мембрани, хоча і на невелику відстань. У постсинаптичній мембрані є не лише хемозбудливі, але й електрозбудливі канали. Тому перехід локального постсинаптичного потенціалу в потенціал дії відбувається у самій постсинаптичній мембрані.
Сумація. Одним із варіантів сумації ЗПСП, що призводить до виникнення ПД, є часова сумація. Вона зумовлена тривалістю існування ЗПСП на постсинаптичній мембрані і виникає у разі дії повторних аферентних імпульсів в одному синапсі. Якщо до пресинаптичної мембрани за короткий проміжок часу будуть надходити ПД, то ЗПСП, що розвивається після кожного з них, накладається на попередній і в разі досягнення критичного рівня перейде у ПД. Таке явище можливе у зв'язку з тим, що звичайно по нервовому волокну надходять не поодинокі ПД, а їх групи - патерни ("пачки").
Інший різновид сумації - просторова. Якщо активність виникає одночасно в більше ніж одному синаптичному контакті, то простежується просторова сумація, й активність одного синапсу підсилює активність іншого, наближаючись до критичного рівня деполяризації.
Синаптична затримка. У зв'язку із названим вище механізмом передачі збудження через синапс швидкість проведення у ньому сповільнюється, що зумовлено часом виділення трансмітера з міхурців пресинаптичної мембрани, часом дифузії його через пресинаптичну мембрану у синаптичну щілину, часом його дії на рецептори постсинаптичної мембрани, що називають си-наптичною затримкою. Синаптична затримка в ЦНС складає 0,2-0,5 мс.