

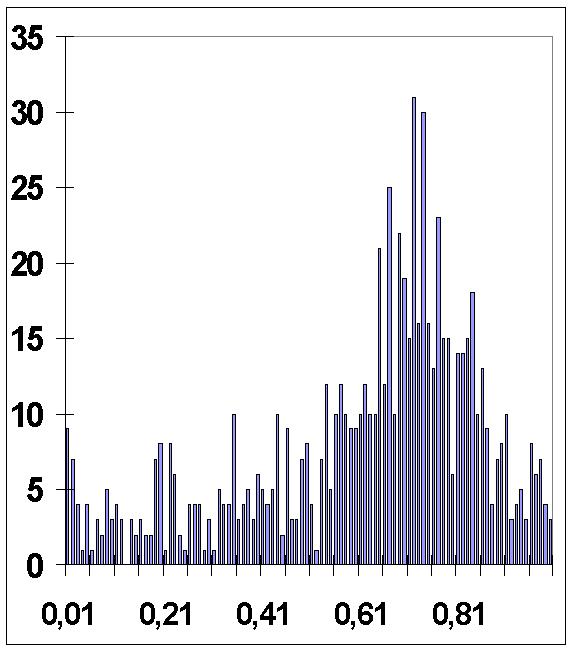
26.1.Рис 32.Гистограмма дробных частей
в![]() сех
времен деления клеток нематоды в единицах
-кванта
времени
сех
времен деления клеток нематоды в единицах
-кванта
времени
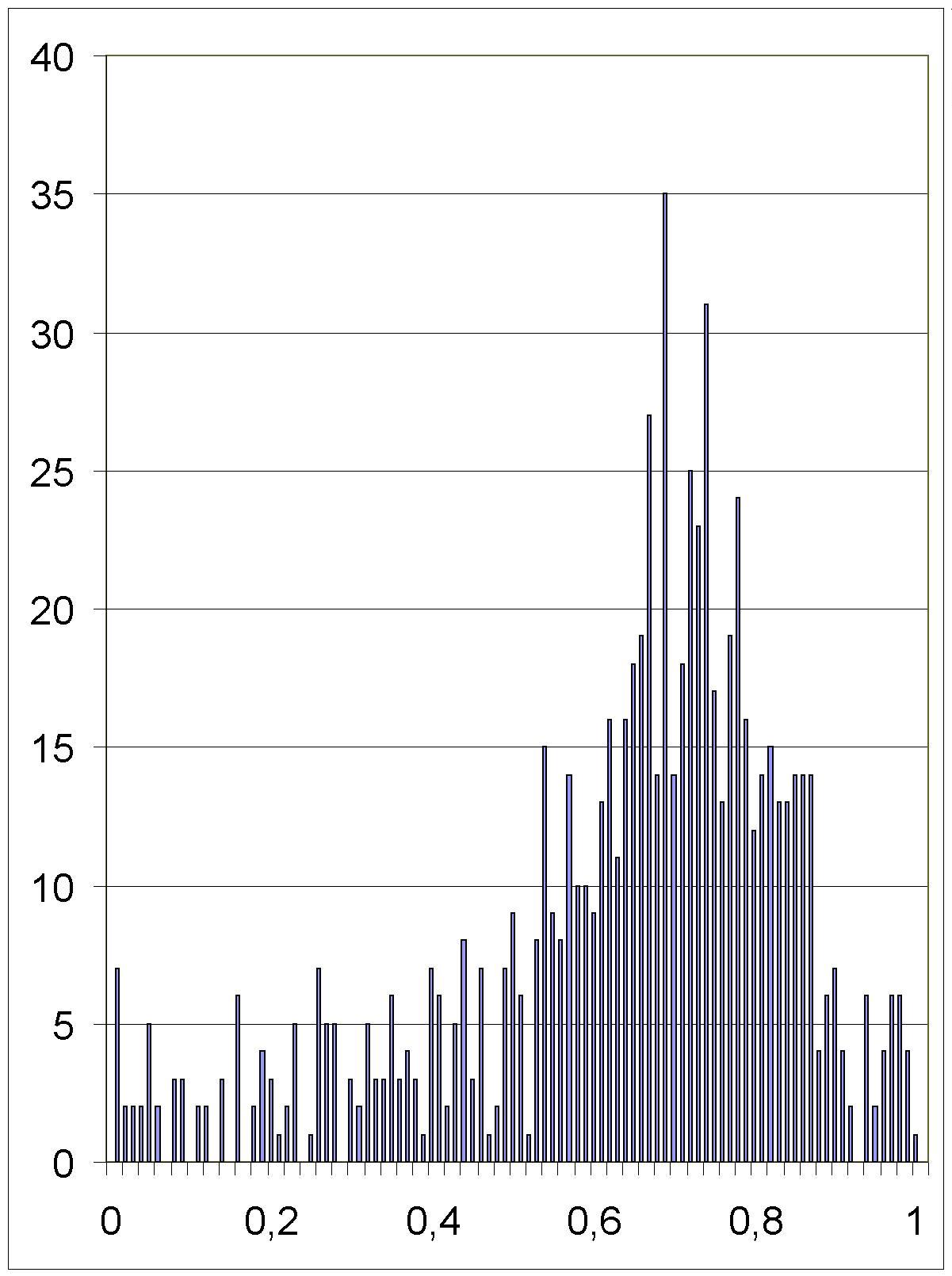
2 6.2.Рис 33. Гистограмма дробных частей последних времен деления клеток нематоды в единицах кванта времени
262Таблица № 18.Для гистограммы дробных частей всех времен деления клеток нематод в единицах
к ванта времени
Рубр. |
Част. |
Рубр. |
Част. |
Рубр. |
Част. |
Рубр. |
Част. |
0,01 |
9 |
0,26 |
4 |
0,51 |
4 |
0,76 |
13 |
0,02 |
7 |
0,27 |
4 |
0,52 |
1 |
0,77 |
23 |
0,03 |
4 |
0,28 |
4 |
0,53 |
7 |
0,78 |
15 |
0,04 |
1 |
0,29 |
1 |
0,54 |
12 |
0,79 |
15 |
0,05 |
4 |
0,3 |
3 |
0,55 |
5 |
0,8 |
6 |
0,06 |
1 |
0,31 |
1 |
0,56 |
10 |
0,81 |
14 |
0,07 |
3 |
0,32 |
5 |
0,57 |
12 |
0,82 |
14 |
0,08 |
2 |
0,33 |
4 |
0,58 |
10 |
0,83 |
15 |
0,09 |
5 |
0,34 |
4 |
0,59 |
9 |
0,84 |
18 |
0,1 |
3 |
0,35 |
10 |
0,6 |
9 |
0,85 |
10 |
0,11 |
4 |
0,36 |
3 |
0,61 |
10 |
0,86 |
13 |
0,12 |
3 |
0,37 |
4 |
0,62 |
12 |
0,87 |
9 |
0,13 |
0 |
0,38 |
5 |
0,63 |
10 |
0,88 |
4 |
0,14 |
3 |
0,39 |
3 |
0,64 |
10 |
0,89 |
7 |
0,15 |
2 |
0,4 |
6 |
0,65 |
21 |
0,9 |
8 |
0,16 |
3 |
0,41 |
5 |
0,66 |
12 |
0,91 |
10 |
0,17 |
2 |
0,42 |
4 |
0,67 |
25 |
0,92 |
3 |
0,18 |
2 |
0,43 |
5 |
0,68 |
10 |
0,93 |
4 |
0,19 |
7 |
0,44 |
10 |
0,69 |
22 |
0,94 |
5 |
0,2 |
8 |
0,45 |
2 |
0,7 |
19 |
0,95 |
3 |
0,21 |
1 |
0,46 |
9 |
0,71 |
15 |
0,96 |
8 |
0,22 |
8 |
0,47 |
3 |
0,72 |
31 |
0,97 |
6 |
0,23 |
6 |
0,48 |
3 |
0,73 |
16 |
0,98 |
7 |
0,24 |
2 |
0,49 |
7 |
0,74 |
30 |
0,99 |
4 |
0,25 |
1 |
0,5 |
8 |
0,75 |
16 |
Еще |
3 |
271.
Р ис.34. Кумулятивные скорости накопления всех событий деления клеток (соб./мин.)к полному времени от оплодотворения.(«грубые» волны митозов)

272.Таблица 19. Исходные данные и кумуля-тивные скорости накопления событий деления клеток (соб./мин.).
n соб. |
tn-врем. соб. |
n/ tn |
n соб. |
tn- врем. соб. |
n/ tn |
n соб. |
tn- врем. соб. |
n/ tn |
2 |
50 |
0,04 |
29 |
152 |
0,19079 |
56 |
182,5 |
0,30685 |
3 |
68,1 |
0,04405 |
30 |
152,1 |
0,19724 |
57 |
182,5 |
0,31233 |
4 |
68,1 |
0,05874 |
31 |
152,6 |
0,20315 |
58 |
182,5 |
0,31781 |
5 |
84,1 |
0,05945 |
32 |
152,6 |
0,2097 |
59 |
182,6 |
0,32311 |
6 |
84,5 |
0,07101 |
33 |
152,7 |
0,21611 |
60 |
182,6 |
0,32859 |
7 |
89,4 |
0,0783 |
34 |
152,8 |
0,22251 |
61 |
182,7 |
0,33388 |
8 |
93,9 |
0,0852 |
35 |
152,8 |
0,22906 |
62 |
182,8 |
0,33917 |
9 |
101,4 |
0,08876 |
36 |
152,8 |
0,2356 |
63 |
182,9 |
0,34445 |
10 |
102,5 |
0,09756 |
37 |
152,9 |
0,24199 |
64 |
182,9 |
0,34992 |
11 |
102,7 |
0,10711 |
38 |
153 |
0,24837 |
65 |
182,9 |
0,35539 |
12 |
103 |
0,1165 |
39 |
153 |
0,2549 |
66 |
182,9 |
0,36085 |
13 |
110,7 |
0,11743 |
40 |
153,3 |
0,26093 |
67 |
183 |
0,36612 |
14 |
113,1 |
0,12378 |
41 |
153,4 |
0,26728 |
68 |
183 |
0,37158 |
15 |
118,6 |
0,12648 |
42 |
153,5 |
0,27362 |
69 |
183 |
0,37705 |
16 |
123,4 |
0,12966 |
43 |
153,6 |
0,27995 |
70 |
183,2 |
0,3821 |
17 |
124 |
0,1371 |
44 |
154,2 |
0,28534 |
71 |
183,2 |
0,38755 |
18 |
124,4 |
0,14469 |
45 |
158,1 |
0,28463 |
72 |
183,2 |
0,39301 |
19 |
124,5 |
0,15261 |
46 |
160,4 |
0,28678 |
73 |
183,3 |
0,39825 |
20 |
124,6 |
0,16051 |
47 |
163,5 |
0,28746 |
74 |
183,3 |
0,40371 |
21 |
124,6 |
0,16854 |
48 |
164,5 |
0,29179 |
75 |
183,4 |
0,40894 |
22 |
124,7 |
0,17642 |
49 |
165,9 |
0,29536 |
76 |
183,5 |
0,41417 |
23 |
124,8 |
0,18429 |
50 |
166,6 |
0,30012 |
77 |
183,7 |
0,41916 |
24 |
127 |
0,18898 |
51 |
166,8 |
0,30576 |
78 |
183,9 |
0,42414 |
25 |
136,9 |
0,18262 |
52 |
181,8 |
0,28603 |
79 |
184,4 |
0,42842 |
26 |
137,1 |
0,18964 |
53 |
182,3 |
0,29073 |
80 |
184,4 |
0,43384 |
27 |
147,8 |
0,18268 |
54 |
182,3 |
0,29622 |
81 |
184,4 |
0,43926 |
28 |
148,9 |
0,18805 |
55 |
182,4 |
0,30154 |
82 |
185,1 |
0,443 |