

34.9...........Последовательность
а.к. ГЕМОГЛОБИНА
34.9.1. Общая характеристика гемоглобина
Гемоглобины
- это белки эритроцитов крови живот-ных.
Гемоглобины всех позвоночных, за
исключением круглоротых, представляют
собой тетромерные моле-кулы, состоящие
из четырех пептидных цепей. Гемо-глобин
(гемоглобин А) содержит две
-цепи
и две
![]() -цепи.
Гемоглобин плода (гемоглобин F)
содержит вместо
-цепей
-цепи.
Гемоглобин плода (гемоглобин F)
содержит вместо
-цепей
![]() -цепи
и еще есть редкий тип гемог-лобина (А2)
взрослого человека, содержащего вместо
-цепей
-цепи
и еще есть редкий тип гемог-лобина (А2)
взрослого человека, содержащего вместо
-цепей
![]() -цепи.
Всего, таким образом, имеется четы-ре
гомологичные цепочки типа
,
,
,
длинною около 140-150 аминокислотных
остатков.
-цепи.
Всего, таким образом, имеется четы-ре
гомологичные цепочки типа
,
,
,
длинною около 140-150 аминокислотных
остатков.
Разбиение гомологичной части аминокислот-
ных цепей гемоглобина человека
В таблице. 31А. представлены 4-ре последователь-ности цепей гемоглобина человека ( , , , ).
Таблица. 31А.
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
12 |
13 |
14 |
15 |
V |
- |
L |
S |
P |
A |
D |
K |
T |
N |
V |
K |
A |
A |
W |
V |
H |
L |
T |
P |
E |
E |
K |
S |
A |
V |
T |
A |
L |
W |
G |
H |
F |
T |
E |
E |
D |
K |
A |
T |
I |
T |
S |
L |
W |
V |
H |
L |
T |
P |
E |
E |
K |
T |
A |
V |
N |
A |
L |
W |



1 |
17 |
18 |
19 |
20 |
21 |
22 |
23 |
24 |
25 |
2 |
2 7 |
28 |
29 |
30 |
G |
K |
V |
G |
A |
H |
A |
G |
E |
Y |
G |
F |
E |
A |
L; |
G |
K |
V |
D |
- |
- |
V |
D |
E |
V |
G |
G |
E |
A |
L |
G |
K |
V |
D |
A |
H |
V |
E |
D |
A |
G |
G |
E |
T |
L |
G |
K |
V |
D |
A |
H |
V |
D |
A |
V |
G |
G |
E |
A |
L |
 6
6 6
6

3 |
32 |
33 |
34 |
35 |
36 |
3 7 |
38 |
39 |
4 0 |
41 |
42 |
4 |
44 |
45 |
E |
R |
M |
F |
L |
S |
F |
P |
T |
T |
K |
T |
Y |
F |
P |
G |
R |
L |
L |
V |
V |
Y |
P |
W |
T |
E |
R |
F |
F |
E |
G |
R |
L |
L |
V |
V |
Y |
P |
W |
T |
D |
R |
F |
F |
E |
G |
R |
L |
L |
V |
V |
Y |
P |
W |
T |
E |
R |
F |
F |
E |
 1
1 3
3
46 |
47 |
48 |
49 |
50 |
51 |
52 |
53 |
54 |
55 |
56 |
57 |
58 |
59 |
60 |
H |
P |
- |
D |
L |
S |
H |
P |
D |
- |
- |
- |
G |
S |
A |
S |
P |
G |
D |
L |
S |
T |
A |
S |
A |
V |
M |
G |
D |
P |
S |
P |
G |
D |
L |
S |
S |
A |
S |
A |
I |
M |
G |
D |
P |
S |
P |
G |
D |
L |
S |
S |
V |
D |
A |
M |
M |
G |
D |
P |

61 |
62 |
63 |
6 |
65 |
66 |
67 |
68 |
69 |
70 |
71 |
72 |
73 |
7 |
75 |
E |
V |
L |
G |
H |
G |
L |
L |
V |
A |
D |
A |
L |
T |
N |
K |
V |
L |
A |
H |
G |
L |
L |
V |
L |
G |
A |
P |
S |
D |
K |
V |
L |
A |
H |
G |
L |
L |
V |
L |
T |
S |
L |
G |
D |
K |
V |
L |
A |
H |
G |
L |
L |
V |
L |
G |
A |
L |
S |
D |
 4
4 4
4
76 |
77 |
78 |
79 |
80 |
81 |
82 |
83 |
84 |
85 |
86 |
87 |
88 |
89 |
90 |
A |
V |
A |
H |
V |
D |
D |
M |
P |
D |
A |
L |
S |
A |
L |
G |
L |
A |
H |
L |
D |
D |
L |
K |
G |
T |
P |
A |
T |
L |
A |
I |
G |
H |
L |
D |
D |
L |
K |
G |
T |
P |
A |
E |
L |
G |
L |
A |
H |
L |
D |
D |
L |
K |
G |
T |
P |
S |
Q |
L |

Таблица.31А,. продолжение
91 |
92 |
93 |
94 |
95 |
96 |
97 |
98 |
99 |
100 |
101 |
102 |
103 |
104 |
105 |
S |
D |
L |
H |
A |
H |
K |
L |
R |
V |
D |
P |
V |
D |
F |
S |
E |
L |
H |
C |
D |
K |
L |
H |
V |
D |
P |
E |
N |
F |
S |
E |
L |
H |
C |
D |
K |
L |
H |
V |
D |
P |
E |
N |
K |
S |
E |
L |
H |
C |
D |
K |
L |
H |
V |
D |
P |
E |
N |
F |



106 |
107 |
108 |
109 |
110 |
111 |
112 |
113 |
114 |
115 |
116 |
117 |
118 |
119 |
120 |
K |
L |
L |
S |
H |
C |
L |
L |
V |
T |
L |
A |
A |
H |
L |
R |
L |
L |
G |
D |
V |
L |
V |
C |
V |
L |
A |
H |
H |
P |
R |
L |
L |
G |
D |
V |
L |
V |
T |
V |
L |
A |
I |
H |
P |
R |
L |
L |
G |
D |
V |
L |
V |
C |
V |
L |
A |
R |
W |
P |


121 |
122 |
123 |
124 |
125 |
126 |
127 |
128 |
129 |
130 |
131 |
132 |
133 |
134 |
135 |
P |
A |
E |
F |
T |
P |
A |
V |
H |
A |
S |
L |
D |
K |
P |
G |
K |
E |
F |
T |
P |
P |
V |
Q |
A |
A |
Y |
Q |
K |
V |
G |
K |
E |
F |
T |
E |
P |
V |
Q |
S |
Y |
Y |
Q |
M |
V |
G |
K |
E |
F |
T |
Q |
M |
V |
Q |
A |
A |
Y |
Q |
K |
V |


-
136
137
1
 38
38139
140
141
1 42
143
144
145
146
147
148
L
A
S
V
S
T
V
L
T
S
L
Y
R
V
A
E
V
A
D
A
L
A
H
L
Y
H
V
T
E
V
A
S
A
L
S
S
R
Y
H
V
A
E
V
A
D
A
L
A
H
L
Y
H
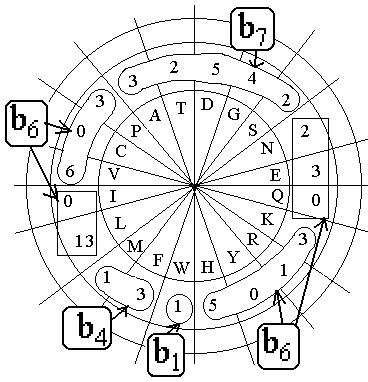
Рис.47Д
Консервативная часть составила 48 аминокис-лот и к ней добавлены 5 аминокислот консерва-тивных в 3-х цепях, а в четвертой-просто отсут-ствует. Кроме того, есть 3 позиции, в которых сочетаются близкие друг к другу аспаргиновая и глутаминовая кислоты и аспаргин и аланин. в парах D-N, D-E. Добавлена позиция №43 для комплектности группы b4. Вместе 48+5+3+1 дает 57 аминокислот. Представление 57 аминокислот по типам и их разбиение на bs даны на Рис. 47Д.
.34.9.2. ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Видно, что группа b7=16 выделена по-прежнему среди малых аминокислот, а группа b1 такая же, что и у консервативной части цитохромов С по шести цитохромам, т.е. b1 - W-аминокислотный остаток. При этом в 4-х смешанных и не вполне консервативных позициях считаем главной D-аминокислоту, а не N,E. Выделены две девятки (по G,S,N,E и по H,Y,R,H). Группа b4 слилась с двумя девятками из b6 =36 : 9+9+4=22, среди C, V, I, L, M, F- аминокислот.