

33.2.1. Общая характеристика фермента
Каталитическая активность этого РНК-фермента была открыта в 1981-1982 гг. в одноклеточном организме Tetrahymena thermophilia, относящегося к типу прос-тейших[25]. Часть РНК, интрон-фермент, может катализировать своё вырезание и сплайсинг РНК, в результате которых из неё выщепляется небольшой фрагмент-интрон.
 Пространственная
структура рибосомного РНК-фермента
принципиальна важна для каталити-ческой
активности этой молекулы. Она
поддержи-вается,
Пространственная
структура рибосомного РНК-фермента
принципиальна важна для каталити-ческой
активности этой молекулы. Она
поддержи-вается,
Рис. 43а
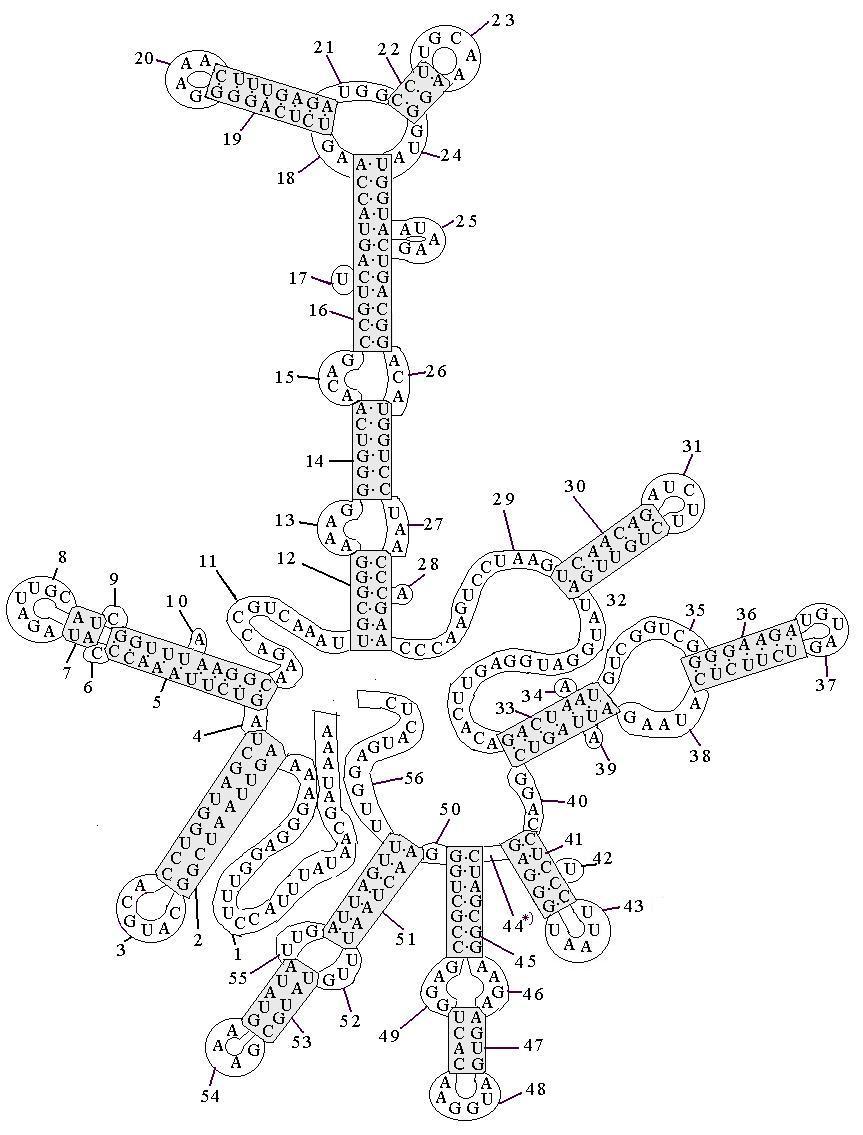
в частности, благодаря способности одноцепочечной РНК сворачиваться, образуя короткие двухцепочечные участки. РНК- это полимер, образованный нуклеатидами четырех видов A, G,C и U. Пары A с U, G с C комплемен-тарны, т.е. образуют устойчивую пару, соединяясь водо-родными связями. (G и U тоже могут спариваться, хотя в этом случае связь гораздо слабее). Полностью структура РНК-фермента тетрахимены представлена на рис. 43а. Участки, где присутствуют подряд только пары или только одиночные аминокислотные остатки пронуме-рованы нами. Участок обозначенный у нас под №44 (44*) выделен самим автором [25] как отдельный, но в нем не был указан ни один нуклеотид.
33.2.2 Разбиение рнк-фермента
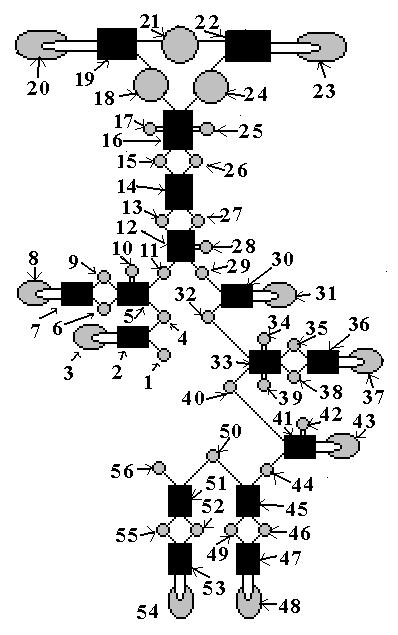 В молекуле есть
участки, образующие комплиментарные
соединения, и участки, не имеющие парных
соединений. Они чере-дуются и пронумеро-ванны
на упрощенном рисунке - схеме фер-мента
(рис.43б.) от 1 до 56. Число 56 подразу-мевает
собой поиск разбиения вида «1+3+36+16» или
вида «4+36+16».
В молекуле есть
участки, образующие комплиментарные
соединения, и участки, не имеющие парных
соединений. Они чере-дуются и пронумеро-ванны
на упрощенном рисунке - схеме фер-мента
(рис.43б.) от 1 до 56. Число 56 подразу-мевает
собой поиск разбиения вида «1+3+36+16» или
вида «4+36+16».
Рис.43б. Разбиение
РНК-фермента.
Элемент №25 выделен тем, что это единственный элемент типа дуги, присоединенной не с торца к двойному участку, а с боку. Его относим к группе b1=1.
Элементы №№18, 21, 24 выделены среди всех тем, что они образуют уникальное кольцо. Их считаем остатком группы b4=4, редуцировавшей до трех.
Дугообразных элементов, замкнутых каждый на торец своего двойного участка, у нас девять : №№3, 8, 20, 23, 31, 37, 43, 48, 54. Их считаем 9-кой из группы b6=36.Двойных участков у нас 16: №№2, 5, 7, 12, 14, 16, 19,22, 30, 33, 36, 41, 45, 47,51,53. Их относим к группе b7=16.
Имеем еще девятнадцать промежуточных между двойными элементами (№№ 4, 6, 9, 11, 13, 15, 26, 27, 29, 32, 35, 38, 40, 44, 46, 49, 50, 52, 55) и шесть одиночных боковых элементов (№№ 10, 17, 28, 34, 39, 42) и два концевых (№№ 1, 56). Всего 27, недостающих в группе b6=36 до 36-ти элементов.
Совместное рассмотрение пар нуклеотидов как целого, интересно тем, что встречается нам также в п.. 6.3., при обсуждении структуры транспортных тРНК. Здесь выступают как равноправные структурные элементы, составленные из одиночных нуклеотидов и их пар, а там (при рассмотрении тРНК, с. 299) равноправны сами одиночные нуклеотиды и их пары.
33.3. Церамид гликосфинголипидов
33.3.1. Общая характеристика
ГЛИКОСФИНГОЛИПИДОВ
В гликолипидах две составные части - углеводный компонент и липидный. Гликосфинголипиды [26]- это особый подкласс гликолипидов, содержащий особый липид сфингозин. Гликосфинголипиды есть во всех клетках животных и некоторых растительных клетках. Располагаясь в клеточной мембранне, молекулы гликосфинголипидов различными путями регулируют взаимодействие клетки с ее окружением. Например, гликосфинголипиды служат маркерами (т.е. они различны на клетках из разных органов и тканей) и таким образом участвуют в процессах узна-вания и контактов клеток. Антигены групп крови и многие другие антигены, которые модифицируются или аномально экспрессируются при развитии злока-чественных опухолей, являются гликосфинголипида-ми. Нервная ткань, и в частности, мозг, весьма богаты гликосфинголипидами, и долгое время наиболее ин-тенсивно этот класс веществ изучали именно нейро-биологи и нейрохимики.
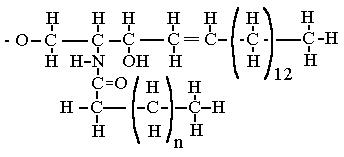
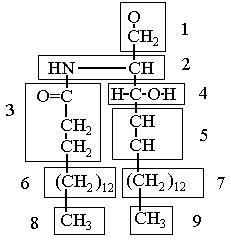
Рис. 44а. Структура цера-
мидного одновалентного радикала, который присо-единяется к цепочке остат-ков сахаров.
Рис.44б. Разбиение радикала церамида на блоки атомов. Блоки пронумерованны от 1 до 9.
Информационная часть гликосфинголипида представ-лена цепочкой простых сахаров, консервативная - липидом, который называется церамидом. Строение как церамида, так и углеводородных цепей может сильно варьировать, и [26] известно уже более 130 различных гликосфинголипидов. Церамид образуется
соединением аминокислоты серина с жирной кислотой с образованием спирта сфингозина с последующим присоединением второй молекулы жирной кислоты. В спирте сфингозина всегда 18 атомов С в цепи. Разнообразие молекул церамида достигается вариацией длины углеводородного остатка 2-й жирной кислоты. Наиболее часто встречается длина 2-го углеродного хвоста с n=13. (см. рис. стр. 358 [27]) Двойных связей в углеродном скелете в хвостах, по-видимому, нет, кроме обозначенных на рис.44а.