

13. Комментарии к разбиению
НА ЭКЛОНЫ
В 1988 году две ветви биологии - популяционная био-логия и генетика индивидуального развития - внезапно нашли общие точки соприкосновения благодаря весьма оригинальной концепции Л. Басса [22] по поводу природы естественного отбора :
«..сам индивидуальный организм - считает Басс (цитата по Л.В. Белоусову [23] ) - в единственном числе, представляя собой химеру из клонов, осуществляет выживание, отбор самих этих клонов, и результатом подобного отбора и будет сам индивидуум, инди-видуальность». Здесь подчеркнем, что именно этой идее решили придерживаться и мы при таком отборе вариантов математического описания динамики раз-вития нематоды по эклонам (элементарным клонам), как в этой главе. Противоположным взглядом на орга-низм является его представление исключительно целостным и согласованным во всех своих частях. В пункте 17 мы не пользуемся никакими эклонами и описываем организм эмбриона нематоды цельным, привлекая для этого решения уравнений дифференцировки для старших s>7, вместо того чтобы многократно использовать один и тот же набор решений при s<8. Последний взгляд нам нравиться больше, а первый-скорее историчен. В нем должным образом выделены первые несколько групп - 55 пар нейронов, 55 клеток, дифференцирующихся после вылупления. Остальные менее наглядны и неодноз-начны.
Ниже (стр.124-129) дана в качестве примера, одна из 16 таблиц [9] (для 55 нейронов и их родословных), со- держащая последовательные времена делений в кле-точных линиях.
Также приведены 14 рисунков для фрагментов всего древа развития нематоды, касающихся каждый одного эклона. В первом выпуске [53] они уже были, но мы приводим их опять.
Повороты ветвей на этих рисунках относительно вертикальных отрезков клеточных линий инвариан-тны(-револьверная симметрия), что приводит лишь к формальному несовпадению клеточных ветвей друг с другом и не всегда совпадают с тем, что были на ис-ходных рисунках [2,].Эта процедура способствует улуч-шению выделению групп bs и bsi .
Таблица 11.1
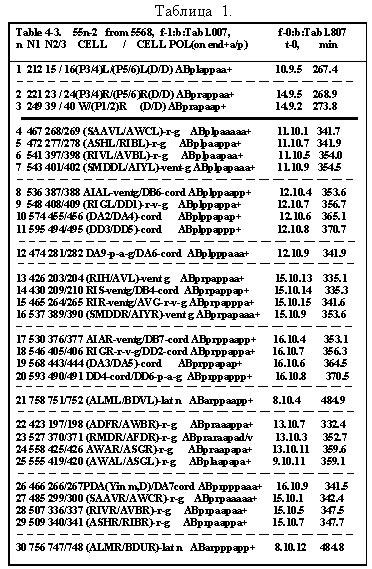
Таблица 11.2.
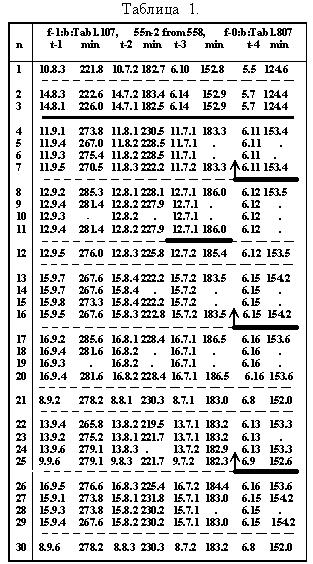
Таблица 11.3
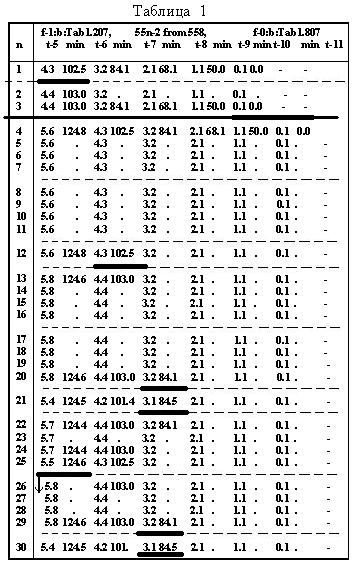
Таблица 11.4
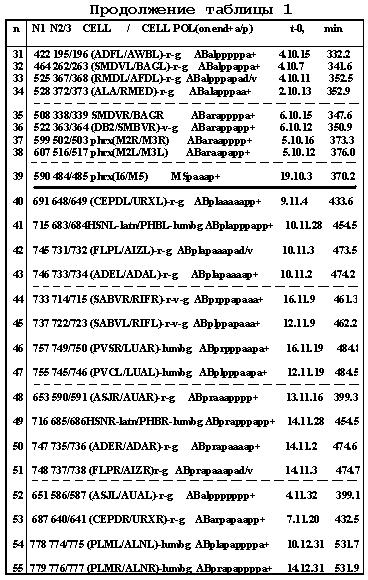
Таблица 11.5
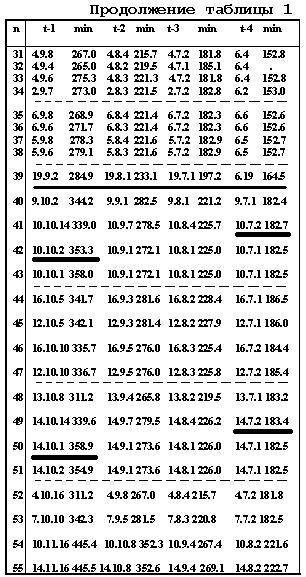
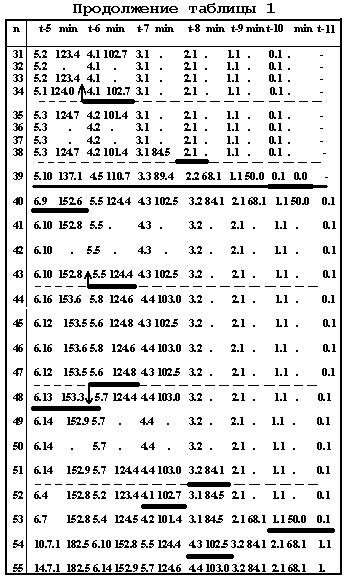 Таблица
11.6
Таблица
11.6
14(16-Ть)клонов всех поклеточных делений
ДЕРЕВА РАЗВИТИЯ ДО ВЫЛУПЛЕНИЯ (784 событий)
С ПРЕДСКАЗАННЫМИ ЧИСЛЕННОСТЯМИ
ЭЛЕМЕНТАРНЫХ КЛОНОВ
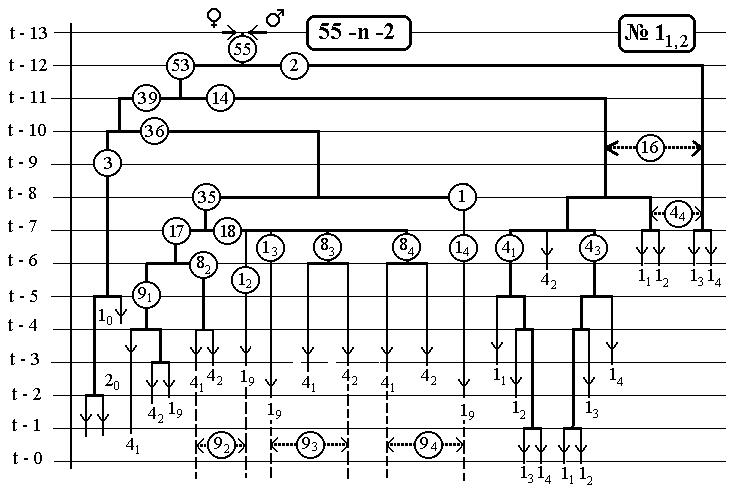
РИС.20 (верхний), РИС.21 (нижний)
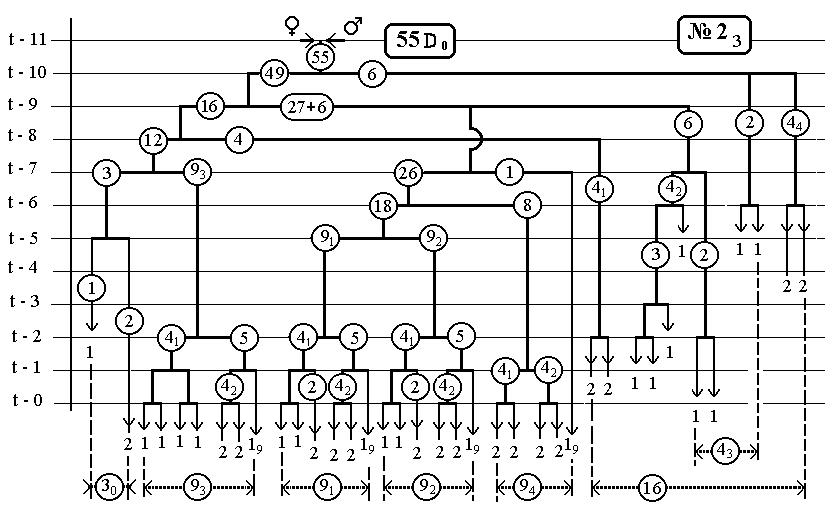
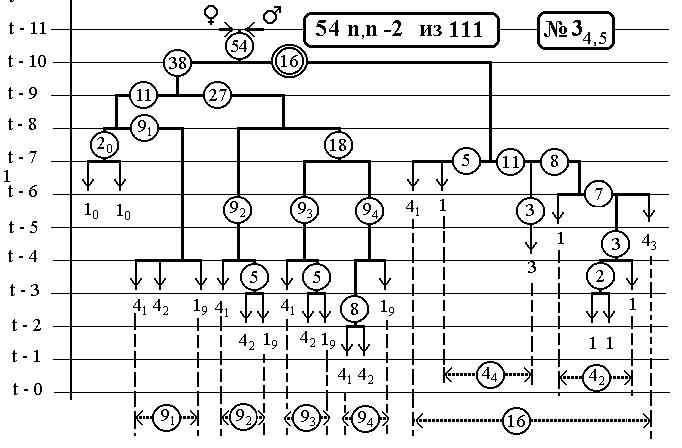


РИС.22 (верхний), РИС.23 (нижний)
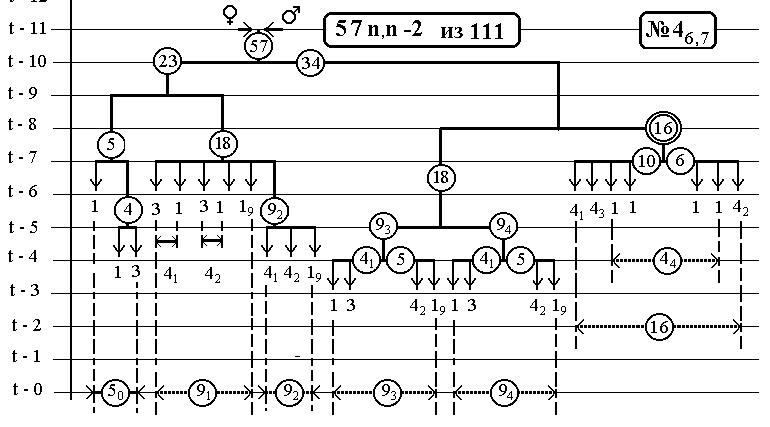

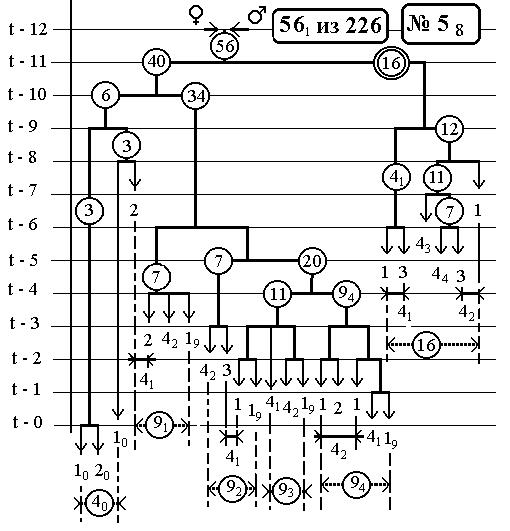
РИС.24 (верхний), РИС.25 (нижний)
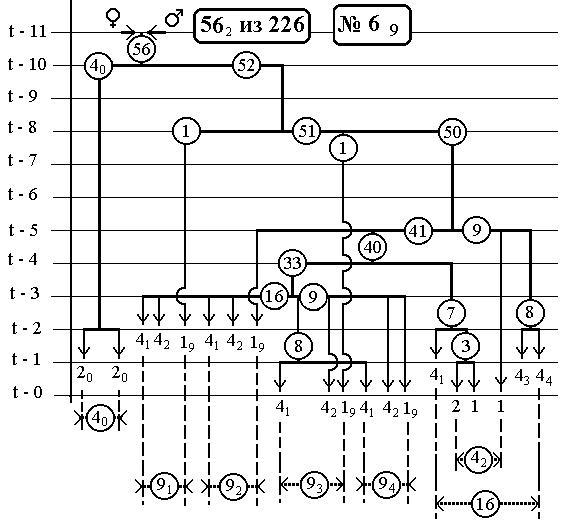
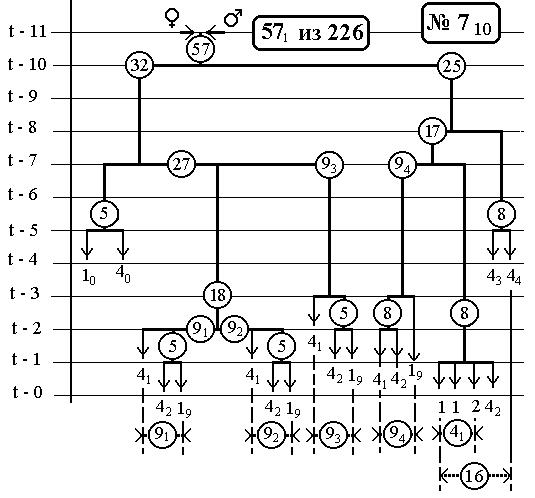
РИС.26 (верхний), РИС.27 (нижний)
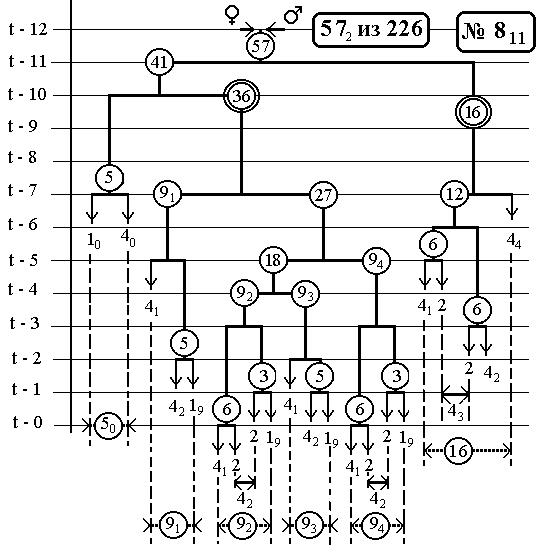
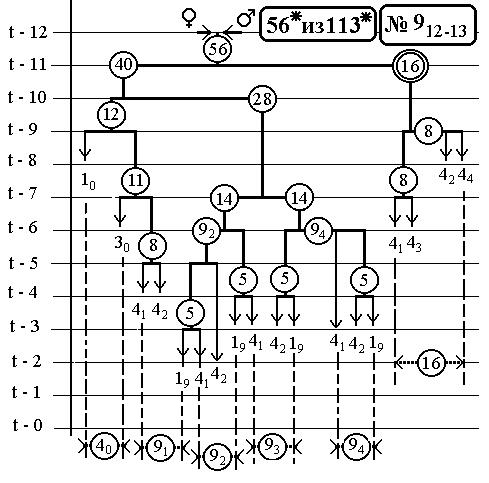
РИС.28 (верхний), РИС.29 (нижний)
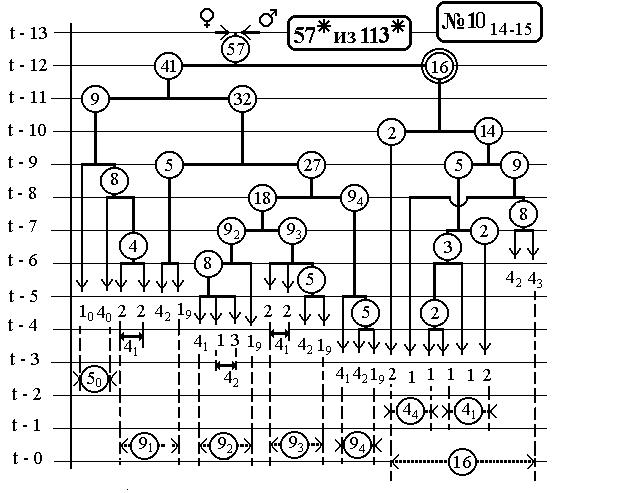
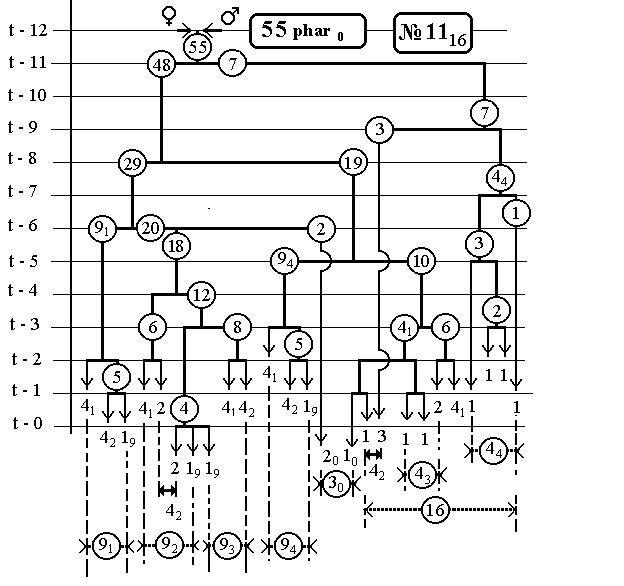
РИС.30.