

8. Эмбриональное развитие c.Elegans.
8.1.Актуальность поиска «кванта вре-
МЕНИ» ИЛИ МИКРОРИТМА В ОБЩЕЙ
ДИНАМИКЕ
По общепринятой логике «квант» (-4 ) это нечто, далее не дробимое(не считая здесь вынужденно введен-ной 4-ки) Но эта величина(«квант») призвана наиболее адекватным способом описывать сложные динамичес-кие явления, их особенности достаточно крупного раз-мера. Отметим, что здесь мы намеренно ввели более мелкую единицу времени- ,умножив её на 4, пос-кольку х 4 часто встречается в приложениях.
На данном предварительном этапе представляется естественным «напрямую» воспользоваться моделью типа ДТА-21, выделяющей интервал (12, 13-20, 21) 4 как область усиленной дифференцировки.

Рис.3. Гистограмма 564 дифференцированных
клеток,
появляющиеся за![]() 800
мин.(784-786)
800
мин.(784-786)
На представленном выше рисунке дана гистограм-ма времен появления 564 дифференцированных клеток на интервале[0-786]мин.(т.е. не делящихся
далее в эмбриональном развитии и не умирающих клеток до вылупления нематоды C. elegans; для самца и гермафродита -объединенная выборка, рубрика данной гистограммы –5 мин., общий диапазон – 800 мин.) Указанных данных на первых порах должно хватить для грубой или интегральной оценки величины .На интервале [260;500] мин.(внутрен-ней части полного интервала [0-786]мин.) появляются 554 клетки из 564–т.е. приблизительно 98,23%.Первое весьма грубое значение для средней величины даёт нам дробь 500/21=23,8095, откуда следует: (500/21)/4=5,95238095 м., где 21-смоделированное чис-ло стадий согласно ДМЯ, явно обнаруживаемое по гистограмме рис.3.
Несколько ниже, используя не интегральную информацию о моментах деления клеток, а выборку из 55 из всех 784 известных нам момента времени, мы сможем вычислить значительно более точное значение в фибоначчиевом представлении :
![]()
=(55 + 5)/10=5,72360679774998.. м.
Используя эту значительно более точную информа-цию, вводя более грубую шкалу времени со 140 -приб-лизительно-рубриками, мы сможем начать непосредс-твенно визуально наблюдать «квантовые» явления, тем самым значительно, расширив диапазон действия
новой единицы времени .Для начала предс-тавим все 784 события в этой расширенной шкале:
Рис.4
НАБЛЮДЕННЫЕ НА ОПЫТЕ СОБЫТИЯ ДЕЛЕНИЯ И
АПОПТОЗА КЛЕТОК ПРИ ЭМБРИОНАЛЬНОМ РАЗВИ-
ТИИ НЕМАТОДЫ C.Elegans.
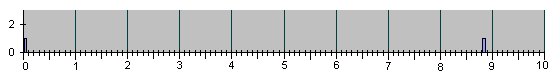
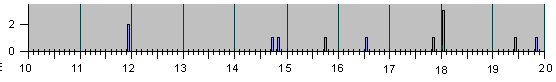
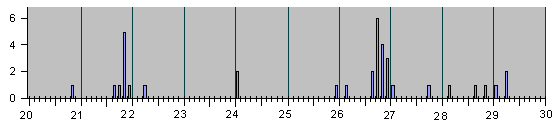
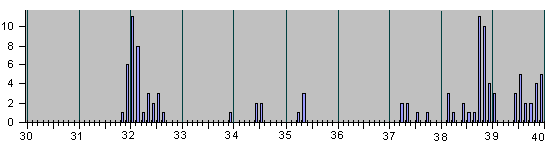
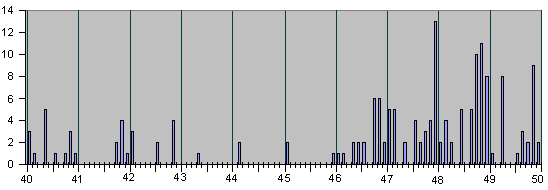
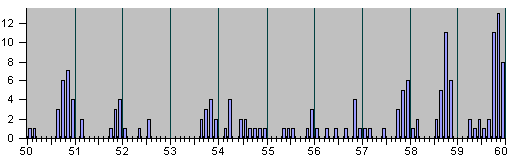
Рис. 4, продолжение
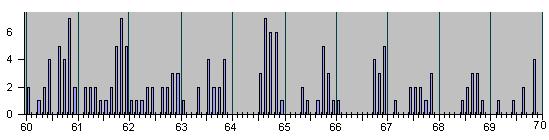
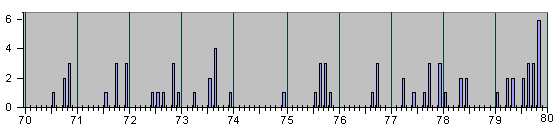
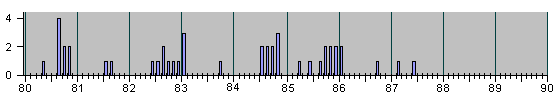
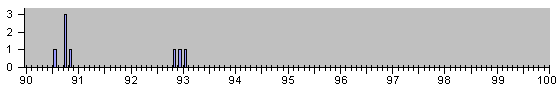
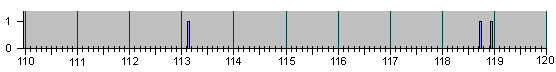
Для ориентации читателя в последующем ниже мы представляем серию гистограм в более сжа-тых шкалах разных масштабов :
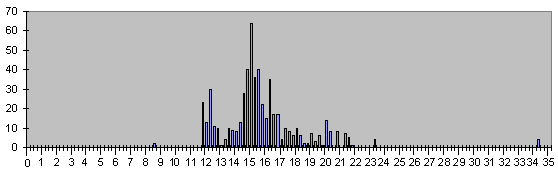
Рис.5, Гистограмма времен появления дифференцированных клеток клеточного древа развития С.Elegans с исправленным по эмпирическому микроритму значением
4 =22,89442719 минуты.
Теперь посмотрим на поведение различных типов событий эмбрионального развития относите-льно модельного ритма с тем же 4 шагом :
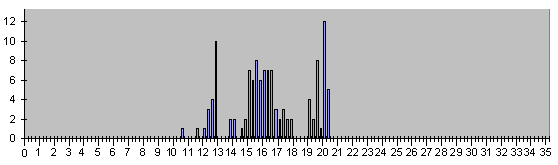
Рис. 6. Гистограмма времен появления уми-рающих в эмбриональном развитии клеток.
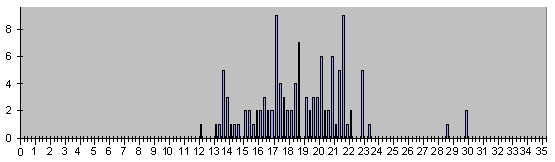
Рис. 7. Гистограмма времен смертей умира-
ющих в эмбриональном развитии клеток.
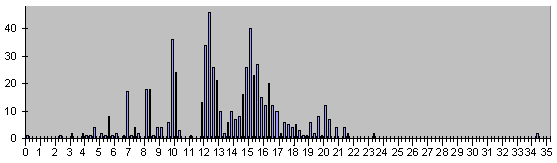
Рис.8. Гистограмма всех событий, кроме смертей .
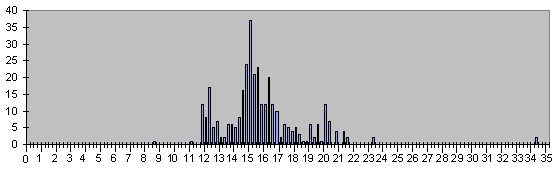
Рис. 9. Гистограмма времен 342 клеточных делений, за которые появляются 564 диффе-ренцированные клетки, этот рис. подобен рис. 5.
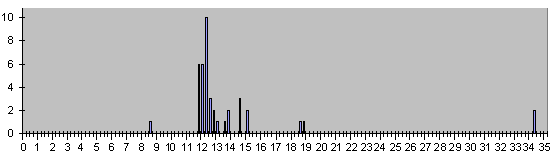
Рис. 10. Гистограмма времен появления 59 клеток, которые делятся в постэмбриональ-ном развитии нематоды С.Elegans.
Для них «4 » несколько меньше прежнего.