

19. Расширение модели до шести
кварков
b8 = 36 = 91+92+ 41+42+43 +44 +11+12 и в ней:
91 = (ubb,dbb,sbb,bU,bD,bS,UBB,DBB,SBB);
92 = (uub,ddb,ssb,uB,dB,sB,UUB,DDB,SSB);
41= (usb,ucb,USB,UCB); 42= (dsb,dcb,DSB,DCB);
43= (udb,scb,UDB,SCB); 44= (cbb,ccb,CBB,CCB);
11 = (bC); 12 = (cB).
b9 = 36 = 91+92+93+ 41+11+12+13+14+15 и в ней:
91 = (utt,dtt,stt,tU,tD,tS,UTT,DTT,STT);
92 = (uut,ddt,sst,uT,dT,sT,UUT,DDT,SST);
93 = (ust,uct,UST,UCT,tC,dst,dct,DST,DCT);
41= (udt,sct,UDT,SDT); 11 = (cbt); 12 = (cct); 13 = (cT); 14 = (CBT);
15 = (CCT).
b10 = 16 = 41+42+11+12+13+14+15+16+ 17+18 и в ней:
41= (bbb,ttt,BBB,TTT); 42= (bbt,btt,BBT,BTT);
11 = (ubt); 12 = (sbt);13 = (dbt); 14 = (cbt);
15 = (UBT); 16 = (SBT); 17 = (DBT); 18 = (CBT).
-144-
Частицы для не попавших в эти два списка сочетаний-(bB) и ( tT) - открыты и представляют собой отдельные острова “стабильности” наподобие двух островов ста-бильности сверхтяжелых ядер по нашей схеме, а (bT),(tB) еще не открыты.
Поясним, что в качестве решений b8 , b9 И b10 взяты варианты, отличные от решений, найденных по схеме В.Серпинского [7,33, Л-1].
20. Пары нуклеозидов в
ДВОЙНЫХ ЦЕПОЧКАХ ДНК И РНК
В двойной спирали ДНК существуют пары (Т,А) и (Ц,Г) нуклеотидов. В “пальцах” РНК ещё две пары (У,А) и (Ц,Г). При считывании РНК с ДНК образуется ещё че-тыре пары (Т,А) и (Ц,Г) (А,У) и (Г,Ц). В последних четы-рёх парах первым указан аминокислотный остаток нук-леотида из ДНК, вторым - из РНК. Также в “пальцах” РНК встречается нестандартная пара (У,Г).
Таблица 7. Численности составляющих пары нуклеозидов частей
№пары |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
поли-нуклеотид |
ДНК |
ДНК |
РНК |
РНК |
ДНК |
ДНК |
ДНК |
ДНК |
РНК |
|
ф-14 |
ф-14 |
ф-15 |
ф-15 |
ф-14 |
ф-14 |
ф-14 |
ф-14 |
ф-15 |
остатки аминокислот |
Т 14 А 15 |
Ц 12 Г 15 |
У 11 А 14 |
Ц 12 А 15 |
Т 14 А 14 |
А 14 У 11 |
Ц 12 Г 15 |
Г 15 Ц 12 |
У 11 Г 15 |
сахарофо-сфатный остов |
ф-14 |
ф-14 |
ф-15 |
ф-15 |
ф-15 |
ф-15 |
ф-15 |
ф-15 |
ф-15 |
поли-нуклеотид |
ДНК |
ДНК |
РНК |
РНК |
РНК |
РНК |
РНК |
РНК |
РНК |
сумма |
56 |
55 |
55 |
57 |
57 |
54 |
56 |
56 |
56 |
|
|
|
|
|
|
|
|
|
|
 сахарофо-сфатный
остов
сахарофо-сфатный
остов
-145-
Всего получается 9 широко распространённых пар. Численности атомов по отдельным составляющим час-тям и в сумме в парах нуклеозидов (нуклеотиды без фос-фатных групп) внутри двойных цепочек полинуклео-тидов указаны в таблице №4.
Видим, что все суммарные численности атомов в парах нуклеозидов лежат в пределах от 54 до 57, что попадает в искомый диапазон 52-57. Интересно, что если бы в паре №3 вместо У стоял остаток Т, то сумма стала бы равной 58 и вышла бы за интересующий нас диапазон.
Общяя структура для пар нуклеозидов дана на Рис. 19.
Радикалы R1 и R6- либо “-Н”, либо “-ОН”; R2 либо “-СН”, либо “-С-СН3”; R3 - содержит 4 атома ( RН2 и О) во всех парах кроме №9 ( в паре №9 два атома O ); R4 содержит три атома (Н,NН) с разными положениями водородной связи во всех парах, кроме №9 (в №9 четыре атома (NН и НN); R5 содержит либо один (-Н), либо три (-NН2).
В следующей таблице четыре девятки из группы b6 =36 составлены из атомов сахаров, из которых исклю-чены 4 водорода (первые две 9-ки) и консервативной части пар аминокислотных остатков, включая R4 цели-ком для №№1-8.Для №9 один из водородов в R4 при этом не считается, в связи с их переизбытком в этом радикале по сравнению с другими парами, отходя в группу b7 . После рисунка, чтобы разобраться с составом групп b1,4 и b7 ,составим таблицу №8, в которых даны радикалы R1-R6 для всех пар нуклеозидов. Для удобства каждый атом в паре пронумерован подряд вверху слева. По таблице видно, что в состав b7 кроме 8 выделенных кружками на рис. 19.
-146-
Рис.19 Общая структура нуклеозидных пар.
Фосфатные группы оставлены.
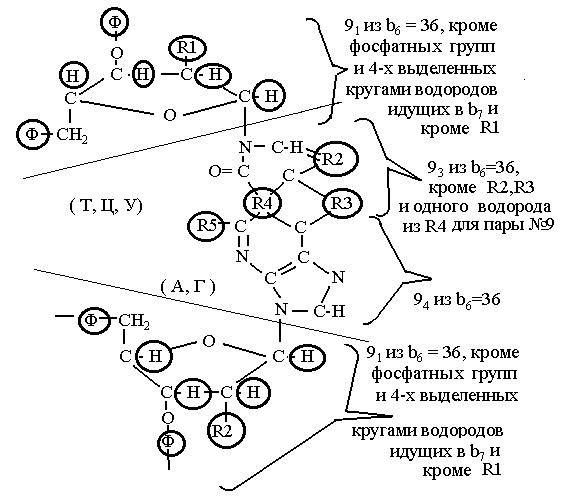
водородов в остатках сахаров и R3 входят ещё водород- только другие водороды.
-147-
Таблица №8. Состав радикалов R1-R6 и выделение групп b1 , b4 и b7 .
№пары |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
R1 |
1Н |
1Н |
1О,2Н |
1О,2Н |
1Н |
1О,2Н |
1Н |
1О,2Н |
1О,2Н |
R2 |
2С,3С 4.5.6Н3 |
2С,3Н |
3С,4Н |
3С,4Н |
2С,3С 4.5.6Н3 |
3С,4Н |
2С,3Н |
3С,4Н |
3С,4Н |
R3 |
7О,8N 9,10Н2 |
4,5Н2 6N, 7О |
5О,6N 7,8Н2 |
5,6Н2 7N, 8О |
7О,8N 9,10Н2 |
5О,6N 7,8Н2 |
4,5Н2 6N, 7О |
5,6Н2 7N, 8О |
5О,6О |
R4 |
11N, 12Н, 13N |
8N, 9Н, 10N |
9N, 10Н, 11N |
9N, 10Н, 11N |
11N, 12Н, 13N |
9N, 10Н, 11N |
8N, 9Н, 10N |
9N, 10Н, 11N |
7N, 8Н, 9Н, 10N |
R5 |
14Н |
11N, 12,13Н2 |
12Н |
12N, 13,14Н2 |
14Н |
12Н |
11N, 12,13Н2 |
12N, 13,13Н2 |
11N, 12,13Н2 |
R6 |
15Н |
14Н |
13О, 14Н |
15О, 16Н |
15О, 16Н |
13О, 14Н |
14О, 15Н |
15О, 16Н |
14О, 15Н |
b7 |
R3, 4Н, 5Н, 6Н, 14Н |
R3, 1Н, 14Н, 12Н, 13Н |
R3, 2Н, 14Н, 4Н, 12Н |
R3, 2Н, 16Н, 13Н, 14Н |
R3, 14Н, 4Н, 5Н, 6Н |
R3, 2Н, 14Н, 4Н, 12Н |
R3, 1Н, 3Н, 12Н, 13Н |
R3, 2Н, 16Н, 13Н, 14Н |
R3, 9Н, 2Н, 12Н, 13Н, 15Н |
b4 |
1Н, 15Н, 2С, 3С |
2С 3Н |
1О, 13О, 3С |
1О, 8О, 3С |
1Н, 16Н 2С, 3С |
1О, 13О, 3С |
3С |
1О, 4Н, 3С |
1О, 14О, 3С |
b1 |
|
11N |
|
12N |
15О |
1О |
11N |
12N |
11N |
Группа b1 состоит из азота радикала -NН3 гуанина ли-бо из одного кислорода -ОН - группы сахарофосфатного остова полипептида .
* * *
-148-
23. СОКРАЩЁННЫЙ СПИСОК ВЫБРАННЫХ
ОБЪЕКТОВ ЕСТЕСТВОЗНАНИЯ, ВНУТРЕННЯЯ
СТРУКТУРА КОТОРЫХ ОПИСЫВАЕТСЯ
РЕШЕНИЯМИ УРАВНЕНИЙ ДИФФЕРЕНЦИРОВКИ
- ПРИМЕНЕНИЯ ДТА”57”
24. Объекты физики, химии и биологии
1. Заполнение нуклонами ядерных оболочек в Fе57 и других устойчивых ядер с весом от 52 до 57 .
2. Совокупность заполнения нуклонами ядерных оболочек 6 атомов трёхвалентного радикала NС2Н2О имеющейся в каждом аминокислотном остатке в полипептидной цепи ( в нём 56 нуклонов в ядрах).
3. РНК - Фермент Tebrahymena thermophila [19, Л-2] . Разбивается на 56 элементов по форме сложения в структуре с “пальцами”.
4. Церамид гликосфинголипидов [20-22, Л-2]. Имеет две части по 55 атомов.
5. Консервативная часть цитохрома-С по 6 ранним по времени определения последовательностям [23, Л-2] имеет 56 аминокислот.
6. Белок гемоглобина . Консервативная часть по 4 цепям у человека имеет 56 аминокислот. Остальные неконсер-вативные 56 аминокислотных позиций также разбива-ются на искомые группы.
7. Ферродоксины типа 1-Fe-S и 8-Fe-S [24-25, Л-2]. Имеют длину в диапазоне от 52 до 57 аминокислотных остат-ков.
8. Антибиотик валиномицин. [26, Л-2] Искусственная молекула облегчающая транспорт иона K+ сквозь мем-браны. Имеет три тождественные группы по 56 атомов.
-149-
9. Витамин В12 .
Имеет часть описывающуюся ДТА”57” при
s=
.
и остаток подпадающий под решения с s=![]() .
.
10. Пара энергетических молекул НАД+Н и НАД+. Работают решения как при s < 8, так и при s > 7.
11. Монактин [27, Л-2].
12. Хлорофилл.
13. Гемоглобин.
14. Коллаген. В его структуре выделяются сходные чере-дующиеся части в 56 и 60 атомов. Служит хорошей иллюстрацией редукции структур некоторых объектов от 60-61 до искомых 52-57 элементов.Такая редукция встречается, например, в молекуле гомеобокса [21, Л-1] и в примере № 23 [21, Л-1].
Некоторые молекулы из каждого ниже приведённого класса (примеры №№15-22) исчерпывающим образом вписываются в идеологию ДТА”57”.
15. Избранные тройки аминокислот в белках (около четверти всех) имеют число атомов от 52 до 56.
16. Многие тРНК имеют нужные нам численности эле-ментов в группах, если считать пары аминокислотных остатков в “пальцах” за отдельные элементы, а остальные аминокислотные остатки считать за один элемент каждый.
17. Многие порфирины (помимо указанных выше гема и хлорофилла) имеют описываемую ДТА”57” структуру. При этом атом в центральном включении это b1 =1, а четыре азота - это b4=4. По четыре углерода рядом с азотами - b7 =16.
18. Стероиды.
19. Фосфолипиды при определённых значениях веса хвостов жирных кислот .
20. Молекула холистерола.
21. Производные простагландинов.
-150-
22. Средний кластер, образованный молекулами воды при 200С содержит 57 молекул [32, Л-2].
25. Объекты биологии развития.
23. Преонкосфера ленточного червя Monieziaxpansa [28, Л-2]. Меняет в процессе развития численность клеток с 60, редуцируясь до 56 [21, Л-1].
24. Некоторые клоны эмбриона нематоды С. Еleqans. Клон парных сестринских нейронов и клон клеток, продолжающих делиться после вылупления из кутику-лы, содержат по 55 элементов (55 пар клеток в первом случае и 55 одиночных - во втором) [7, Л-1].
25. Клеточное дерево глотки С. Еleqans [30, Л-2] имеет 57 последних делений на своих ветвях [12, Л-1]. На группы и подгруппы разбиение идёт по форме ветвей, на котрых находятся эти деления.
26. Клеточное дерево нервной системы асцидии C.Intesti-nalis из [29, Л-2] имеет также 57 последних клеточных де-лений. Разбиение проведено по форме ветвей в [12, Л-1].
27. Всё клеточное
дерево эмбриона С. eleqans описано в [9, Л-2]
с помощью решений ДТА“57” с bs
: s=![]() по форме клеточных ветвей и типу
получающихся готовых клеток.
по форме клеточных ветвей и типу
получающихся готовых клеток.
26(28-36).
НЕКОТОРЫЕ ОПУБЛИКОВАННЫЕ НАМИ РАНЕЕ
ПРИМЕНЕНИЯ АТТРАКТОРА ДTА “21” (№№28-34)
С ЧЁТКИМ ВЫДЕЛЕНИЕМ ИНТЕРВАЛА (12-13,
20-21)*
![]() 0
.
0
.
28. Появление дифференцированных клеток у эмбриона нематоды С. Еleqans ( 0 = 22.94 мин.).
29. Время появления населённых пунктов в геоселитебных системах Подолии ( 0= 75 лет).
30. Появление нейробластов у асцидии ( 0= 1 час).
31. Созревание тимуса у человека ( 0= 1 неделя).
-151-
32.Протекание беременности у человека ( 0= 2 недели )
33. Цикл жёлтого тела у яйцеклетки человека ( 0=день).
34. Половое созревание человека ( 0= 1 год).
Добавим здесь два еще не опубликованных примера.
35. Период половой активности (продуктивность семянников) у нематоды С. Еleqans ( 0= 16 * 22.94 мин ).