

7.4. Более сложная динамика спорообразо-вания у хлореллы .
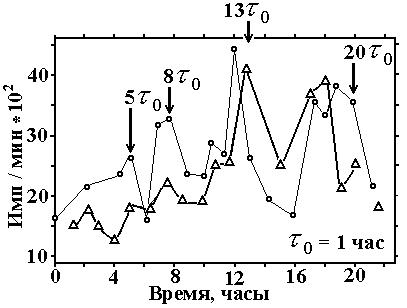 Рис.3
Рис.3
На рис. 3 изображено включение импульсной метки
тимидина - 14С в ДНК клеток штамма ЛАРГ-1 (8-ми
-109-
споровый штамм) в синхронной культуре. = 1 час. s= 5-пик скорости первого синтеза ДНК хлоропласта, s = 8 - пик скорости первого синтеза ДНК, s1, ДНК для 2-х спор, точнее их предшественника, s = 13 - пик скорости второго синтеза ДНК (s2 - уже 4-е набора - предшественников ДНК), s = 19-20 - 3-й (всего 4-й) пик скорости синтеза ДНК для уже всех 8 спор, s3. В среднем по двум приведённым графикам на период в 21 час приходится 7 пиков скорости синтеза ДНК (с учетом "скрытых", выявленных по второй производной от количества ДНК). Отсюда определяется и период процесса S o = 3 час. В этом примере присутствует все главные, предсказываемые ДМЯ особенности: на 5,8, [13-20] шагах и t= 3 . [1, Л-3]
7.5. Краткий перечень других объектов.
Далее, в нашей работе [12, Л-1] приведены примеры для слизневика, динамики инкубации куриных яиц, земно-водных, перевиваемых раковых опухолей (8 - штаммов) , для мышей - динамика накопления 8-ми пищеварите-льных энзимов в эмбриогенезе) с особенностями на 13-21 часах. Остановимся подробнее на слизневике Dictyо-stelium discoideum. На рис. 10 из [12, Л-1] приведена динамика синтеза и появления 189 из 778 всех извес-тных белков (рис.11) у слизневика. Четко определена выделенность 8-го, 12-го и 22 часов. В этом примере мы заинтригованы тем, что у этого животного выделено именно 778 белков. Согласно нашим вариантам (см. п. 1.6.1. [12, Л-1]) и нашему предсказанию здесь на двумерных электрофореграммах не идентифицировано 4-6 белков. Число 778 с точностью 0,8% близко к 784
-110-
(782 - для самца) числу всех событий (делений и смертей клеток, опыт) у гермафродита нематоды. Мы уверены, что преагрегационный период у слизневика - до 10,5 часов - вполне отвечает периоду эмбриогенеза (до вылупления) у нематоды и резонно предполагать, что каждому акту, событию, отражающемуся на формальном виде, структуре дерева развития, отвечает свой белок. Иными словами, этот вывод и лежащие в его основе факты согласуются, как с существованием гомеобокса (см. ниже), так и с основными следствиями из "модели-ядра", с наличием короткой и универсальной (-см. приведенные примеры ) программы для ранних этапов развития - от 1-го до 36-го в единицах , отличных друг от друга для разных видов и таксонов.
* * *
-111-
7.6 Nематоdе c.Elegans -
14(16-Ть)клонов всех поклеточных делений
ДЕРЕВА РАЗВИТИЯ ДО ВЫЛУПЛЕНИЯ (784 событий)
С ПРЕДСКАЗАННЫМИ ЧИСЛЕННОСТЯМИ
ЭЛЕМЕНТАРНЫХ КЛОНОВ
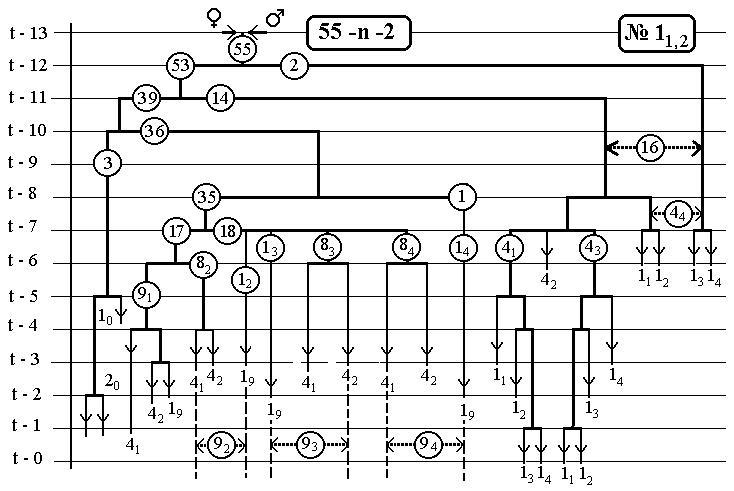
РИС.4 (верхний), РИС.5 (нижний)
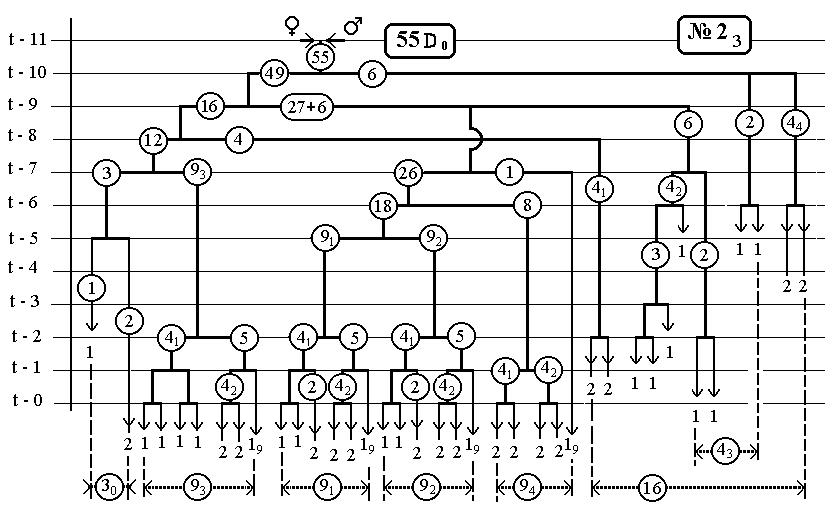
-112-

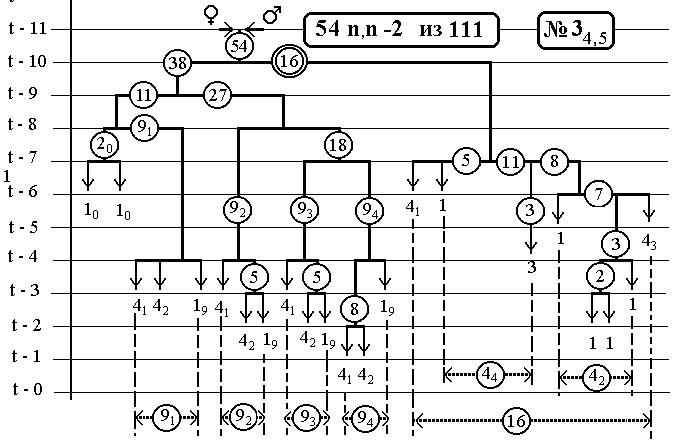

РИС.6 (верхний), РИС.7 (нижний)
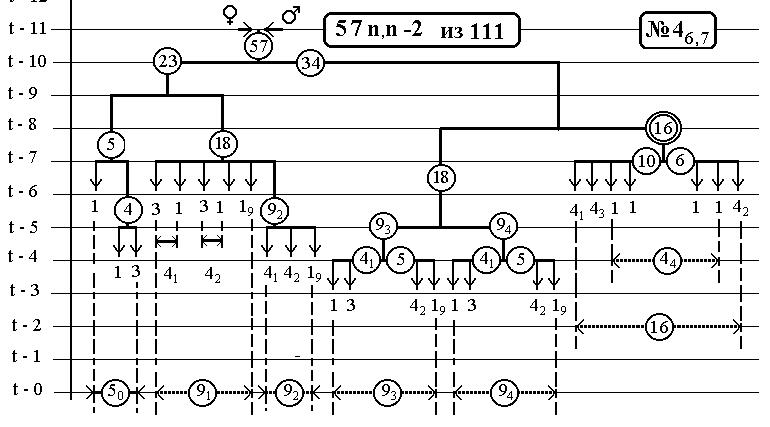

-113-
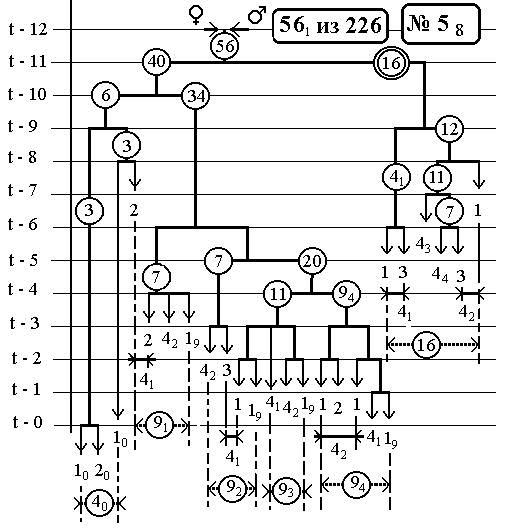
РИС.8 (верхний), РИС.9 (нижний)
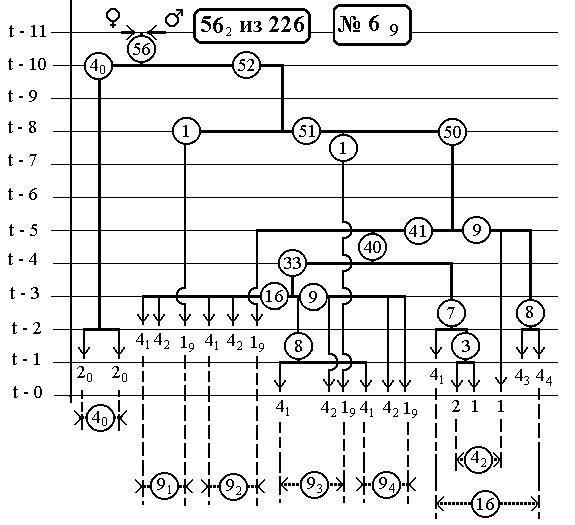
-114-
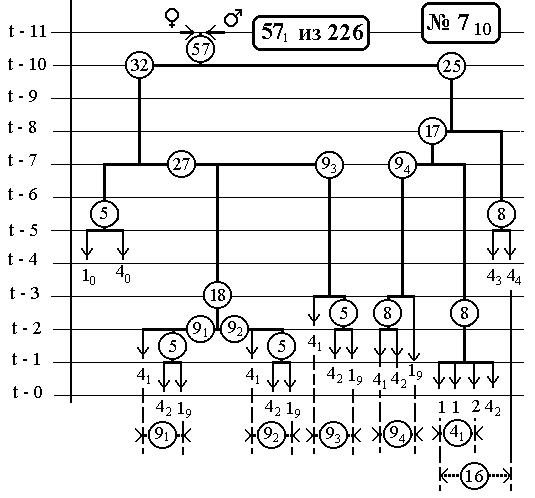
РИС.10 (верхний), РИС.11 (нижний)
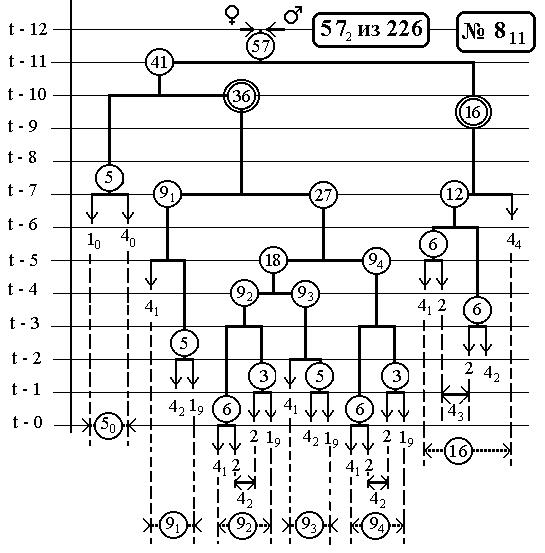
-115-
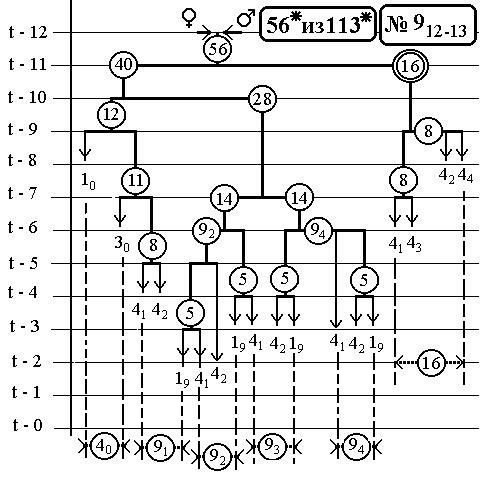
РИС.12 (верхний), РИС.13 (нижний)
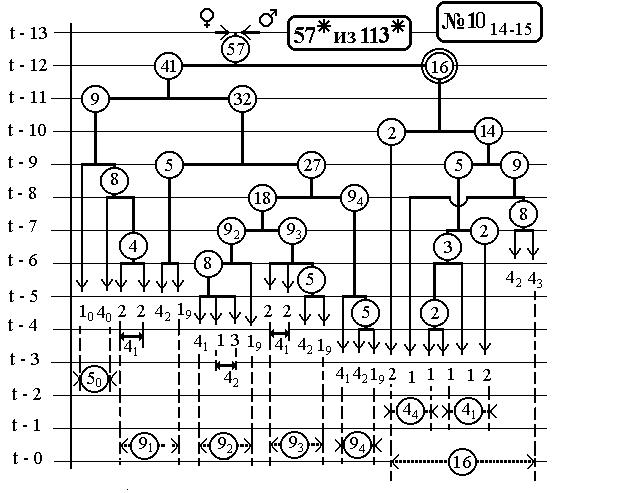
-116-
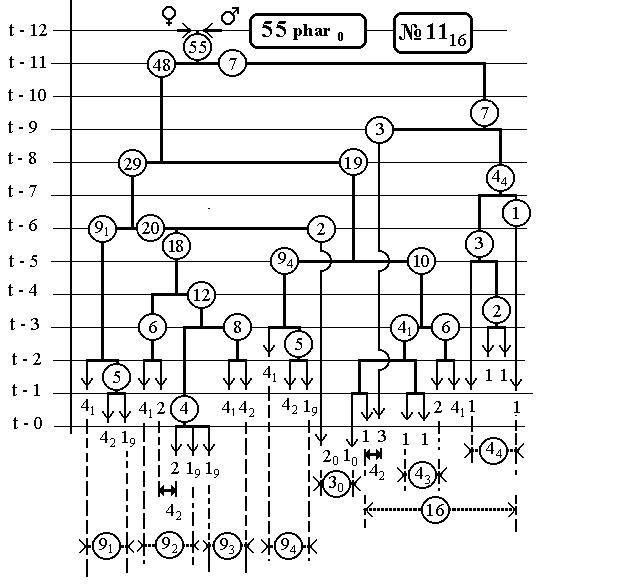
РИС.14.