

3. Структура і склад біомембран
Мембрани складаються з ліпідів трьох класів: фосфоліпіди, гліколіпіди і холестерол. Фосфоліпіди і гліколіпіди (ліпіди з приєднаними до них вуглеводами) складаються з двох довгих гідрофобних вуглеводневих "хвостів", які пов'язані із зарядженою гідрофільній "головою". Холестерол надає мембрані жорсткість, займаючи вільний простір між гідрофобними хвостами ліпідів і не дозволяючи їм згинатися. Тому мембрани з малим вмістом холестеролу більш гнучкі, а з більшим - більш жорсткі й тендітні. Також холестерол служить "стопором", що перешкоджає переміщенню полярних молекул з клітки і в клітку. Важливу частину мембрани складають білки, які пронизують її і відповідають за різноманітні властивості мембран. Їх склад і орієнтація в різних мембранах розрізняються.
Клітинні мембрани часто асиметричні, тобто шари відрізняються за складом ліпідів, перехід окремої молекули з одного шару в інший (так званий фліп-флоп) утруднений.
4. Мембранні органели
Це замкнуті одиночні або пов'язані один з одним ділянки цитоплазми, відділені від гіалоплазми мембранами. До одномембранним органел відносяться ендоплазматична мережа, апарат Гольджі, лізосоми, вакуолі, пероксисоми; до двумембранним - ядро, мітохондрії, пластиди. Будова мембран різних органел відрізняється за складом ліпідів і мембранних білків.
5. Виборча проникність
Клітинні мембрани мають виборчої проникністю: через них повільно дифундують глюкоза, амінокислоти, жирні кислоти, гліцерин і іони, причому самі мембрани певною мірою активно регулюють цей процес - одні речовини пропускають, а інші ні. Існує чотири основних механізми для надходження речовин у клітину або виведення їх з клітини назовні: дифузія, осмос, активний транспорт і екзо-або ендоцитоз. Два перших процесу носять пасивний характер, тобто не потребують витрат енергії; два останні - активні процеси, пов'язані зі споживанням енергії.
Вибора проникність мембрани при пасивному транспорті обумовлена спеціальними каналами - інтегральними білками. Вони пронизують мембрану наскрізь, утворюючи свого роду прохід. Для елементів K, Na і Cl є свої канали. Щодо градієнта концентрації молекули цих елементів рухаються в клітину і з неї. При подразненні канали натрієвих іонів розкриваються, і відбувається різке надходження в клітку іонів натрію. При цьому відбувається дисбаланс мембранного потенціалу. Після чого мембранний потенціал відновлюється. Канали калію завжди відкриті, через них в клітку повільно потрапляють іони калію.
№9.
Прямій поділ клітин (амітоз) та його характеристика.
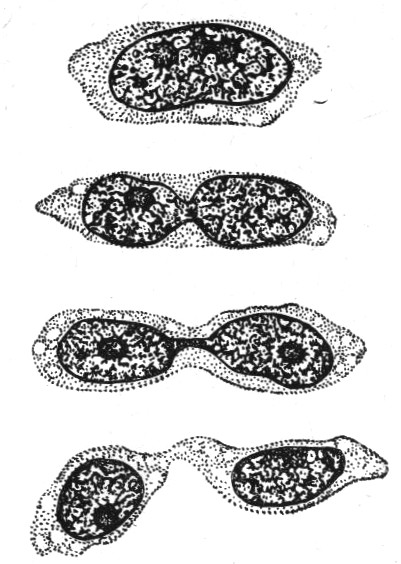
Амітоз (від грец. а— заперечна частка і mitos — нитка) — прямий поділ ядра, який відбувається шляхом перешнуровування ядерної речовини, без утворення хромосом. Явище А. Вперше описав німецький біолог Р.Ремарк (1841). Термін “А” запровадив німецький гістолог В. Флемінг (1882).
Суть амітозу полягає в тому, що ядро, а за ним вміст клітини ділиться на дві частини — дочірні клітини без будь-яких попередніх змін структури органел, в тому числі і ядра.
Причому ядро ділиться на дві частини навіть без попереднього розчинення ядерної оболонки. Відсутнє формування веретена поділу, яке характерне для інших типів поділу.
Після поділу ядра починає ділитися протопласт і вся клітина на дві частини, але в тих випадках, коли спостерігається дроблення ядра на декілька частин, утворюються багатоядерні клітини. При амітозі не відбувається рівномірного розподілу речовини ядра між дочірніми ядрами, тобто не забезпечується їх біологічна рівномірність. Однак утворені клітини не втрачають своєї структурної організації і життєдіяльності.
Довгий час в науці існувала думка, що амітоз — це патологічне явище, яке властиве лише патологічно зміненим клітинам. Проте останні дослідження не підтверджують цієї точки зору. Багатьма дослідженнями (Каролінська, 1951 та ін.) доведено, що амітоз спостерігається і в молодих нормально розвинутих клітинах. Цей тип поділу клітини і ядра спостерігали в клітинах міжвузлів харових водоростей, в клітинах цибулі, традесканції. Крім того, амітоз зустрічається і в спеціалізованих тканинах з високою активністю метаболічних процесів, а саме: в клітинах тапетум мікроспорангіїв, в ендоспермі насіння деяких рослин тощо.
Однак цей тип поділу не зустрічається в клітинах в яких необхідно зберегти повноцінну генетичну інформацію, наприклад, в яйцеклітинах та клітинах зародка. Тому на думку ряду вчених, амітоз не може вважатися повноцінним способом розмноження клітин.
№10.
Поділ соматичних клітин: мітоз та його фази.
Мітоз (непрямий поділ): визначення, фази мітозу та їх характеристика, відкриття мітозу.
Мітоз (непрямий поділ)(від грец. mitos— нитка) — поділ ядра і клітини, при якому з однієї материнської диплоїдної клітини утворюються дві ідентичні дочірні диплоїдні клітини. Розрізняють фази мітозу: профаза, метафаза, анафаза, телофаза. Період між двома поділами називається інтерфазою. Ряд стадій М. Описані Е. Страсбургером на рослинах (1876 - 1879). Детальні дослідження М. Вперше проведені німецькими вченими — ботаніком Е. Страсбургером на рослинах (1876-1879) і гістологом В. Флемінгом на тваринах.
Суть мітозу полягає в тому, що у період підготовки материнської клітини до поділу число хромосом збільшується в два рази. Під час мітозу хромосоми рівномірно розташовуються в двох ядрах дочірніх клітин. Завдяки мітозу форма і число хромосом дочірніх клітин такі ж які в їх попередника — материнської клітини. Звідси чітко видно, що існують біологічні механізми, які забезпечують однакові спадкові властивості материнської клітини та її дочірніх клітин.
Безперервний процес поділу клітини умовно розподіляють на дві окремі фази: період між двома поділами називається інтерфазою, або інтеркінезом.

№11.
Поділ клітин: мейоз та його фази.
Мейоз (редукційний поділ): визначення, І-й і ІІ-й поділи мейозу, фази мейозу та їх характеристика.

Мейоз (редукційний поділ) (від грец. meiosis — зменшення) — один з поділів ядра і клітини, при якому відбувається зменшення кількості хромосом вдвічі, причому з однієї диплоїдної клітини утворюються чотири гаплоїдні. Залежно від місця М. в життєвому циклі організмів розрізняють три типи М.: гаметний, який відбувається в статевих органах і призводить до утворення гамет (у деяких нижчих рослин), зиготний, що здійснюється в зиготі відразу після запліднення (у багатьох водоростей і грибів), споровий або проміжний, який відбувається при спороутворенні (у більшості рослин). М. включає два послідовні поділи: гетеротипний, який супроводжується зменшенням (редукцією) кількості хромосом удвічі і гомеотипний (екваційний), що схожий на мітоз. М. забезпечує рівномірний розподіл генетичного матеріалу в ряді поколінь. М. у тварин вперше описав німецький ботанік Е. Страсбургер (1888), а у рослин вперше описав М. німецький ботанік Е. Страсбургер.
Фази мейотичного поділу подібні до фаз мітотичного поділу. Однак суттєвою різницею між мейозом і мітозом є те, що на початку мейозу гомологічні хромосоми сходяться (кон’югують) попарно і в анафазі ці парні хромосоми розходяться до протилежних полюсів, а при мітозі хромосоми не сходяться попарно, а подвоюються і до полюсів розходяться їх половинки — хроматиди. Внаслідок мейозу число хромосом в клітині зменшується вдвоє, тобто редукує, тому цей поділ називають редукційним поділом. Крім того, весь цикл мітозу відбувається в один поділ клітини, а мейоз завершується за два поділи.
Незважаючи на подібність фаз при мітозі і мейозі, класичне розділення мітозу на стадії не може бути застосоване для описання складних рухів хромосом при мейозі.
№12.
Спадковий апарат людини: будова та характеристика. (ДНК та РНК)
Нуклеїнові кислоти — це складні високомолекулярні біополімери, мономерами яких е нуклеотиди. Вперше їх виявлено в ядрі клітин, звідки й походить назва цих сполук (від лат. нуклеус — ядро). Молекула нуклеотиду складається із залишків нітратної основи, п’ятивуглецевого моносахариду (пентози) і фосфатної кислоти Залежно від виду пентози, що входить до складу нуклеотиду, розрізняють два типи нуклеїнових кислот: дезоксирибонуклеїнову (ДНК) та рибонуклеїнову (РНК). До складу ДНК входить залишок дезоксирибози, а РНК — рибози.
У молекулах ДНК і РНК містяться залишки таких нітратних основ: аденіну (скорочено позначається літерою А), гуаніну (Г), цитозину (Ц). Крім того, до складу ДНК входить залишок тижіну (Т), а РНК -урацилу (У). Отже, до складу молекул ДНК і РНК входить по чотири типи нуклеотидів, які відрізняються за типом нітратної основи.
Нуклеїновим кислотам, як і білкам, притаманна первинна структура — певна послідовність розташування нуклеотидів, а також складніша вторинна і третинна структури, які формуються завдяки водневим зв’язкам, електростатичним та іншим взаємодіям.
Окремі нуклеотиди сполучаються між собою у ланцюг за допомогою особливих «містків» між залишками пентоз двох сусідніх нуклеотидів. Ці «містки» є різновидом міцних ковалентних зв’язків.
Що собою становить структура ДНК? Розшифрування структури ДНК має свою передісторію. 1950 року американський учений Ервін Чаргафф та його колеги, досліджуючи склад ДНК, виявили певні закономірності кількісного вмісту залишків нітратних основ у її молекулі:
- кількість аденінових залишків у будь-якій молекулі ДНК дорівнює числу тимінових (А = Т), а гуані-нових — цитозинових (Г = Ц);
- сума аденінових і гуанінових залишків дорівнює сумі тимінових і цитозинових (А+Г = Т+Ц).
Це відкриття сприяло встановленню просторової структури ДНК і визначенню її ролі в перенесенні спадкової інформації від материнської клітини до дочірньої, від одного покоління організмів до іншого.
1953 року Джеймс Уотсон і Френсіс Крик запропонували модель просторової структури ДНК (мал. 17), правильність якої згодом було підтверджено експериментально. Молекула ДНК складається з двох ланцюгів нуклеотидів, які з’єднуються між собою за допомогою водневих зв’язків. Ці зв’язки виникають між двома нуклеотидами, які ніби доповнюють один одного за розмірами. Встановлено, що залишок аденіну (А) завжди сполучається із залишком тиміну (Т) (між ними виникає два водневі зв’язки), а гуаніну (Г) — із залишком цитозину (Ц) (між ними виникає три водневі зв’язки) (мал. 18). Чітка відповідність нуклеотидів у двох ланцюгах ДНК має назву комплементарність (від лат. комплементум — доповнення).
Відповідно до запропонованої моделі будови ДНК два ланцюги нуклеотидів обвивають один одного, створюючи закручену праворуч спіраль (вторинна структура ДНК). При цьому діаметр спіралі становить приблизно 2 нм (нанометр дорівнює 1*10-6 міліметра).
За певних умов (дія кислот, лугів, високої температури тощо) відбувається процес денатурації ДНК -розривання водневих зв’язків між комплементарними
нітратними основами різних полінуклеотидних ланцюгів. При цьому ДНК повністю або частково розпадається на окремі ланцюги, через що втрачає свою біологічну активність. Денатурована ДНК після припинення дії факторів, які її спричиняють, може поновити свою структуру завдяки відновленню водневих зв’язків між комплементарними нуклеотидами (процес ренатурації ДНК).
Завдяки здатності формувати структури вищих порядків (третинну тощо) молекула ДНК набуває вигляду компактних утворів. Наприклад, довжина молекули ДНК найбільшої хромосоми людини дорівнює приблизно 8 см, але вона укладена таким чином, що міститься в хромосомі завдовжки лише приблизно 5 нм. Це стає можливим завдяки просторовому ущільненню дволанцюгової спіралі ДНК з утворенням тримірної структури — суперспіралі. Це зумовлено взаємодією між ДНК і ядерними білками клітин еукаріотів.
У багатьох прокаріотів, деяких вірусів, а також у мітохондріях і хлоропластах еукаріотів ДНК з білками не взаємодіє і має кільцеву структуру.
Які функції ДНК? Ви вже знаєте, що одиницею спадковості всіх організмів є ген — ділянка молекули ДНК (а у деяких вірусів — РНК). Він несе спадкову інформацію про структуру певного білка або нуклеїнової кислоти (детальніше про організацію спадкового матеріалу різних груп організмів ми розглянемо далі). Саме ДНК зберігає спадкову інформацію в організмі та забезпечує її передачу дочірнім клітинам під час поділу материнської.
Що собою становлять рибонуклеїнові кислоти? Молекули рибонуклеїнових кислот (РНК) мають подібну до ДНК будову, але складаються лише з одного ланцюга {мал. 19). У деяких вірусів трапляються і дволанцюгові РНК. Відомо три основні типи РНК: інформаційна, або матрична (іРНК, або мРНК), транспортна (тРНК) і рибосомна (рРНК). Вони розрізняються місцем розташування в клітині, формою, розмірами та функціями.
Інформаційна РНК є копією певної ділянки молекули ДНК (одного чи кількох генів). Вона переносить спадкову інформацію від ДНК до місця синтезу молекули білка (мал. 20), а також бере безпосередню участь у її збиранні. Частка іРНК становить приблизно 2% загальної кількості РНК клітини. Вторинна і третинна структури іРНК формуються за допомогою водневих зв’язків, електростатичних та інших типів взаємодій. Молекула іРНК відносно нестабільна, вона швидко розпадається на нуклеотиди. Наприклад, у мікроорганізмів іРНК існує усього декілька хвилин, а в клітинах еукаріотів — декілька годин або днів. Транспортна РНК порівняно з інформаційною має менші розміри. її частка становить до 20% загальної кількості РНК у клітині. Вона приєднує до себе амінокислоти і переносить їх до місця синтезу білкової молекули. Кожну амінокислоту транспортує специфічна тРНК. Транспортна РНК має постійну вторинну структуру, яка за формою нагадує листок конюшини (мал. 21). Така просторова структура зумовлена водневими зв’язками між комплементарними нуклеоти-дами. Біля верхівки такого «листка конюшини» розташовані три нуклеотиди, що визначають, яку саме амінокислоту слід транспортувати. Сама амінокислота приєднується за допомогою ковалентного зв’язку до ділянки біля основи молекули тРНК.
Рибосомна РНК становить приблизно 80% загальної кількості РНК у клітині. Вона входить до складу особливих органел клітин усіх типів — рибосом (детальніше про їхню будову та функції ви дізнаєтеся згодом). Взаємодіючи з білками, рРНК виконує структурну функцію і бере певну участь у процесах синтезу білків. Але в передачі спадкової інформації вона участі не бере.
Аденозинтрифосфатна кислота (АТФ) за будовою подібна до нуклеотидів, із яких складаються РНК.
Що собою становить АТФ? Які її функції? Молекула АТФ складається із залишків нітратної основи (аденіну), вуглеводу (рибози) та трьох залишків фосфатної кислоти. АТФ — універсальна сполука. У її високоенергетичних хімічних зв’язках запасається значна кількість енергії. Якщо за участю відповідного ферменту від молекули АТФ відщепляється один залишок фосфатної кислоти, АТФ перетворюється на аденозиндифосфатну кислоту (АДФ) (мал. 22). При цьому звільняється приблизно 42 кДж енергії. Коли ж від молекули АТФ відщепляються два залишки фосфатної кислоти, вона перетворюється на аденозинмонофосфатну кислоту (АМФ). При цьому енергії звільняється вже до 84 кДж. Отже, під час розщеплення молекули АТФ виділяється велика кількість енергії. Вона використовується для синтезу необхідних організму сполук, на підтримання певної температури тіла, забезпечення різних процесів життєдіяльності. Під час утворення молекул АДФ з АМФ та АТФ з АДФ у зв’язках, що виникають між залишками молекул фосфатної кислоти, запасається відповідна кількість енергії. Тому молекула АТФ є універсальним хімічним акумулятором енергії в клітині.