

Вопрос 6
Пировиноградная кислота является продуктом окисления глюкозы. Ее судьба
различна в зависимости от доступности кислорода в клетке. В анаэробных условиях
она восстанавливается до молочной кислоты. В аэробных условиях происходит ее
окислительное декарбоксилирование до уксусной кислоты, переносчиком которой
служит коэнзим А.
Превращение, состоящее из пяти реакций, осуществляется полиферментным
комплексом, прикрепленным к внутренней митохондриальной мембране со стороны
матрикса. В составе комплекса насчитывают 3 фермента и 5 коферментов.
С О С Т А В П И Р У В А Т Д Е Г И Д Р О Г Е Н А З Н О Г О К О М П Л Е К С А
Пируватдегидрогеназа (Е1), ее коферментом является тиаминдифосфат, ката-
лизирует 1-ю реакцию. (витамин В1)
Дигидролипоамид-ацетилтрансфераза (Е2), ее коферментом является липое-
вая кислота, катализирует 2-ю и 3-ю реакции.
Дигидролипоат-дегидрогеназа (Е3), кофермент – ФАД, катализирует 4-ю и 5-ю
реакции.
Помимо указанных коферментов, которые прочно связаны с соответствующими
ферментами, в работе комплекса принимают участие коэнзим А и НАД.
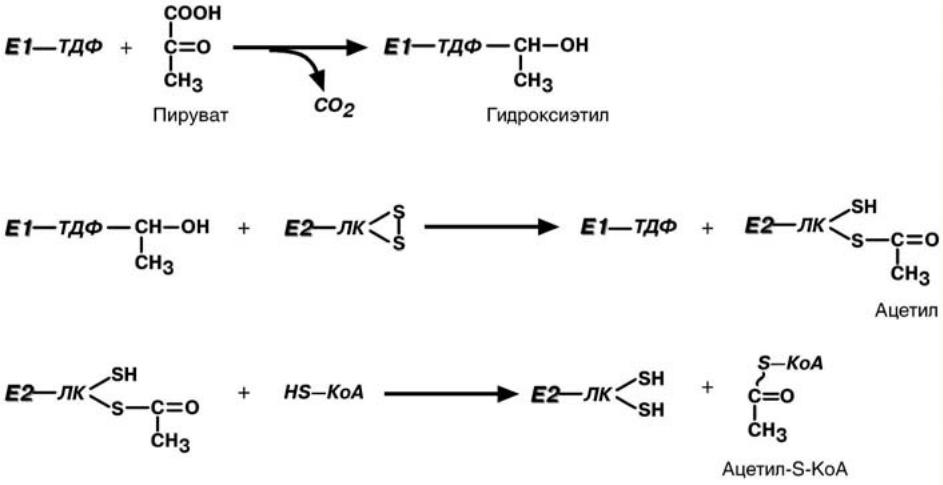
Стадия 1-пируват соединяется с ТДФ в составе Е1 и подвергается декарбоксилированию.
Стадия 2-дигидролипоилтрансацетилаза( Е2) катализирует перенос атомов водорода и ацетильной группы от ТДФ с образованием ацетилтиоэфира липоевой кислоты.
Стадия 3-КоА взаимодействует с ацетильным производным Е2, в результате образуется ацетил-КоА.
Стадия 4- Е3 катализирует перенос атомов водорода от восстановленных липоильных групп на ФАД.
Стадия 5- востанновленный ФАД*Н2 передает водород на НАД+ с образованием НАДН.
Цикл лимонной кислоты(ЦТК)-заключительный этап катаболизма, в котором углерод ацетильного остатка ацетил-КоА окисляется до 2 молекул СО2. Атомы водорода, освобождающиеся в окислительно-востановительных реакциях, под действием дегидрогеназ доставляются в ЦПЭ при участии НАДН+ и ФАДН2, в результате происходят синтез воды и окислительное фосфорилирование АДФ. В условиях организма окисление ацетильного остатка происходит в результате 8 реакций.

Регуляция
В пируватдегидрогеназном комплексе имеются еще 2 вспомогательных регуля-
торных фермента – киназа и фосфатаза, участвующие в фосфорилировании-
дефосфорилировании пируватдегидрогеназы (Е1).
Киназа может активироваться избытком АТФ, НАДН, ацетил-S-КоА. При этом она
фосфорилирует пируватдегидрогеназу, инактивируя ее.
Фосфатаза, активируясь кальцием или инсулином, отщепляет фосфат и активи-
рует пируватдегидрогеназу.
Таким образом, в состоянии покоя при достаточном количестве АТФ пиру-
ватдегидрогеназа неактивна, окисление глюкозы прекращается и она может
использоваться на какие-либо синтетические процессы. Как только клетка
начинает работать, уровень АТФ и НАДН резко снижается, киназная реакция
останавливается, в результате дефосфорилирования пируватдегидрогеназа
активируется, начинается окисление глюкозы.


Пируват + НАД+ + HS-KoA –> Ацетил-КоА + НАДН + Н+ + СO2.
Вопрос 7
Ц И К Л Т Р И К А Р Б О Н О В Ы Х К И С Л О Т
Образующийся ацетил-S-КоА вступает в цикл трикарбоновых кислот (ЦТК, цикл лимонной кислоты, цикл Кребса).
Цикл Кребса протекает в матриксе митохондрий, включает 8 реакций. Это метаболический путь состоит из реакций, в результате которых ацетильный остаток ацетил-КоА окисляется до СО2 и Н2О.Его основная роль заключается в генерации атомов водорода для работы дыхательной цепи, а именно 3-х молекул НАДН и 1-й молекулы ФАДН2. Кроме этого, в ЦТК образуется 1 молекула АТФ; предшественник гема сукцинил-S-КоА; кетокислоты, являющиеся аналогами аминокислот – альфа-кетоглутарат для глутаминовой кислоты, оксалоацетат для аспарагиновой.

Регуляция
1. Главным и основным регулятором ЦТК является оксалоацетат, а точнее его доступность. Наличие оксалоацетата вовлекает в ЦТК ацетил-S-КоА и запускает цикл.
В нормальных условиях в клетке имеется баланс между образованием ацетил-S-КоА (из глюкозы, жирных кислот или аминокислот) и количеством оксалоацетата. Источниками оксалоацетата является синтез из пирувата (анаплеротическая или пополняющая реакция), поступление из фруктовых кислот самого ЦТК (яблочной, лимонной),образование из аспарагиновой кислоты.
Недостаток в клетке глюкозы приводит к сниженному образованию пировиноградной кислоты, что
влечет за собой снижение количества оксалоацетата. Одновременная активация окисления жирных кислот и накопление ацетил-S-КоА запускает резервный путь утилизации ацетильной группы – синтез кетоновых тел. В организме при этом развивается закисление крови (кетоацидоз) с характерной
клинической картиной: слабость, головная боль, сонливость, снижение мышечного тонуса, температуры тела и артериального давления.