43) Окислительное фосфорилирование - главный механизм синтеза атф в
клетке. Представление о хемоосмотической (протондвижущей) теории
Митчелла. Коэффициент Р/О. Пункты сопряжения окисления и
фосфорилирования. Зависимость интенсивности тканевого дыхания в
клетке от концентрации АДФ - дыхательный контроль.
Мех-м окислительного фосфор-я.Теория Митчелла.
Нарушения работы ЦТЭ:
-врождённый
-приобретённый
При наличии врождённого-несовмест-ые с жизнью и новорожденные умирают в течении 1 недели после рождения. Имеется потология-миопатия,наблюдается дефект 3 комплекса дых.цепей. Для коррекции вводят вит.гр К.Благодаря этому обесп. работу дых.цепей в обход повреждённого участка.
Пи наличии приобретенного -впервые в 80-х XX века,у подростков V кустарным способом получили «новый героин». При его изготовлении получили новый продукт-метилфенилтетрогидропиридин-нейротоксин. В кл головного мозга нарушает работу 1 комплекса ферментов дых путей—►болезнь Паркенсонаъ
При работе дых. цепей кроме обр-я эндогенной воды в процессе переноса ёвыд-я Е. Е выд-я на всех участках дыхательных путей,но кол-во выд-ся Е на разных уч разное.Было заметно что кол-во выд-ой Е зависит от редокспотенциала 2х соседних уч-ов дых цепи. Причём чем больше разность тем больше выд Е. Мол-ла V улавливает Е наз АДФ. В результате улавливания(генерирования Е)АДФ—>АТФ. АТФ-энергетическая валюта клетки. Процесс синтеза АТФ из АДФ и неорг-ого фосфата за счёт Е,выд-й при переносе ё дых цепи наз окислительным фосфорилированием.
При ОФ включают 2 процесса:
Экзергоническое окисление восстан-х кофакторов НАДН,ФАДН2 в ходе р- ии тр-а ё
Эндергоническое фосфорилирование АДФ с обр-ем АТФ
Схема окис-ого фосфорилирования:
1 пример: НАДНН*....+1Y202—>НАД* + Н20 +Ag
ФАДН2 +1Y202—>ФАД+Н20 +Ag
2пример: АДФ+НЗР04—>ATФ+H20-Лg(30-40 кДж)
Оба процесса связаны Еки 1 пр-поставления Е,2при-использования Е. Оба процесса идут на внутренней мембране митохондрии.
Было доказано,что такое кол-во Е,выд-я на тех участках д.цепи ,где разность р-п составляет 0,2 V и более.
В дых-ой цепи 1 типа было установлено 3 таких участка:
НАДНН—>ФМН
В^С1
ааЗ—>1\202
В дых цепи 2 типа
В^С1
аа3^1\202
Уч-и дых цепи,где выд-я Е достаточная дл синтеза АТФ наз участками сопряжения или пункты фосфорилирования.
Математически интен-ть процесса можно оценить с помощью коэф.окисл-ого фосф-я К0Ф=Р\0
Р-фосфор,О-кислород
КОФ-величина^ пок-ет какое кол-во неорг-ого фосфата исп-я для синтеза АТФ из АДФ в расчёте на каждый атом потребляемого 02
Для дых цепи 1типа к=3 для дых цепи 2 типа к =2
Мех-м окислиельного фосфорилирования:
гипотеза-химическая
Связь м\у синтезом АТФ и ткан.дых-м осущ-я спец в-ми.
гипотеза-конфармационная
Перенос Е осущ-я в результате конф-ых измен-ий белков,^ уч-ют в этих процессах
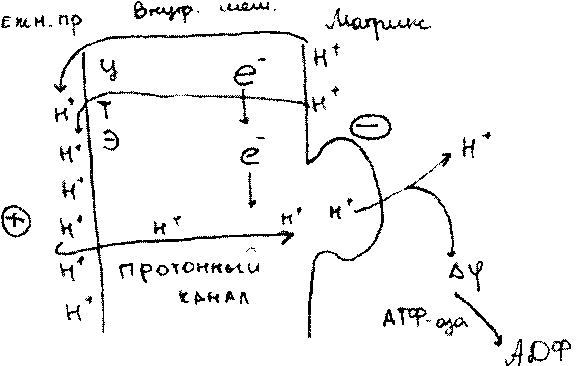
.гипотеза-хемиосмотическая, протон движущая теория,т.Митчела
Согласно т.Митчелла,по ЦТЭ переносятся только ё, V при этом выд-я исп-я для переноса протонов из матрикса в межмембранное пространство митох-ий,т.е дых цепь работает как
протонный насос,V перекачивает протоны(ионы Н)в межмембр-ое пространство. Назад в матрикс протоны вернуться не смогут,т.к. мембрана мит-ий не проницаема.
Тогда они накапливаются на наружной пов-и внутр-ей мембраны и создают протонный градиент(разность концентрации ионов Н матриксе межмембранном пространстве)В рез- те этого наружн. поверхность мит-ой мембраны заряжается «+»,а внутренняя обращённая в матрикс «-». Это приводит к тому что возникает А ц Н * (эл е ктрох и м и чес к и й потенциал) Когда Д|Ш* достигает определённой величины произойдёт сброс протонов ч\з спец-ые протонные каналы в мем-е мит-ий. В составе протонных каналов имеются алигомерные белки(4 стр-ра) V передают др от др ионы Н*,протоны в рез-те движения ч\з протонные каналы происходит активация / Н*-АТФ-аза
При переносе протонов в матриксе происходит резкое падение электрохимического потен-а,меняется Е Ag. Эта Е используется АТФ-азой для синтеза АТФ