

КЛАССИФИКАЦИЯ
УГЛЕВОДОВ (Внелекционный материал для
повторения^.
Углеводы можно определить как альдегидные или
. . j nvy.iiittl VI
(содержащих более одной ОН-группы) спиртов или как соединения, при гидролизе которых образуются эти производные.
Согласно принятой в настоящее время классификации, углеводы подразделяются на три основные группы: моносахариды, олигосахариды и полисахариды.
>
■ жеоды
1СЭДУ р"-
I
Монооаэсарцды
Полисахариды

сашрады и тд.) |
Альдшы
Кетоэы.
,v
I
Г©терогкши- {
шщриды
|
Тшттт*
сахарида




МОНОСАХАРИДЫ
Моносахариды можно рассматривать как производные многоатомных спиртов,
содержащие карбонильную (альдегидную или кетонную) группу. Если карбонильная
группа находится в конце цепи, то моносахарид представляет собой альдегид и
называется альдозой; при любом другом положении этой группы моносахарид является кетоном и называется кетозой.
Простейшие представители моносахаридов - триозы: глицеральдегид и диоксиацетон При окислении первичной спиртовой группы трехатомного спирта - глицерола - образуется глицеральдегид (альдоза), а окисление вторичной спиртовой группы приводит к образованию диоксиацетона (кетоза).
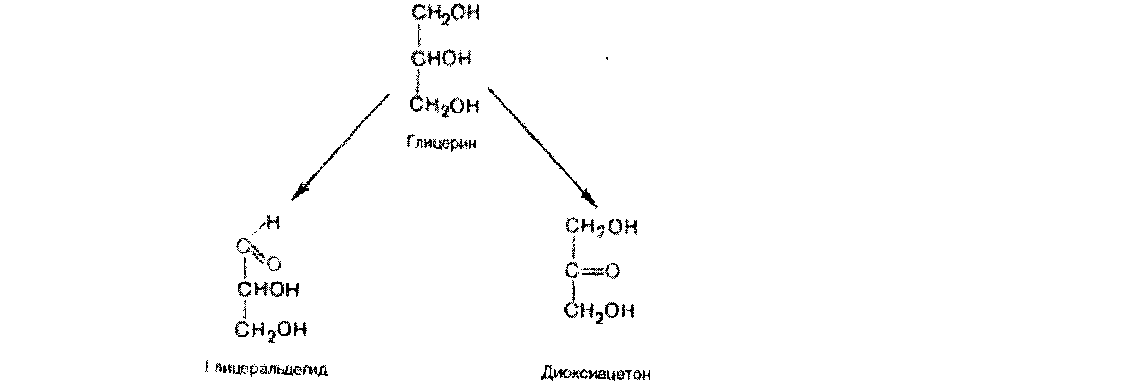
%
содержит асимметричных атомов углерода. Все остальные моносахариды могут существовать в виде различных стереоизомеров.
Общее число стереоизомеров для любого моносахарида выражается формулой N = 2П, где N - число стереоизомеров, ап- число асимметричных атомов углерода. Как отмечалось, глицеральдегид содержит только один асимметричный атом углерода и поэтому может существовать в виде двух различных стереоизомеров.
Изомер глицеральдегида, у которого при проекции модели на плоскость ОН-группа у
„™„„гггт ™Т1ТТГ1™ ^тттг^о^т тл...
асимметричшл'и aiVNLa yi лсрида раишлил^спа с ираиии viupunm, прпплш j_y-
глицеральдегидом, а зеркальное отражение - L-глицераль дегид ом:
f» Vх»
и—о—он но—онн
D-f яйц#раяйй“и# 1-гяиц#раяьд8Гид
Альдогексозы содержат четыре асимметричных атома углерода и могут существовать в виде 16 стереоизомеров (24), представителем которых является, например, глюкоза. Для альдопентоз и альдотетроз число сте-реоизомеров равно соответственно 23 = 8 и 22 = 4.
Все изомеры моносахаридов подразделяются на D- и L-формы (D-и L-конфигурация) по сходству расположения групп атомов у последнего центра асимметрии с расположением групп у D- и L-глицеральдегида. Природные гексозы: глюкоза, фруктоза, манноза и галактоза - принадлежат, как правило, по стереохимической конфигурации к соединениям D-ряда (схема 5.1).
Известно, что природные моносахариды обладают оптической активностью. Способность вращать плоскость поляризованного луча света - одна из важнейших особенностей веществ (в том числе моносахаридов), молекулы которых имеют асимметричный атом углерода или асимметричны в целом. Свойство вращать плоскость поляризованного луча вправо обозначают знаком плюс (+), а в противоположную сторону - знаком минус (-). Так, D-глицеральдегид вращает плоскость поляризованного луча вправо, т. е. D- глицеральдегид является 0(+)-альдотриозой, а L-гли-церальдегид - Ц-)-альдотриозой. Однако направление угла вращения поляризованного луча, которое определяется асимметрией молекулы в целом, заранее непредсказуемо. Моносахариды, относящиеся по стереохимической конфигурации к D-ряду, могут быть левовращающими. Так, обычная форма глюкозы, встречающаяся в природе, является правовращающей, а обычная форма фруктозы - левовращающей.
Циклические (полуацетальные) формы моносахаридов. Любой моносахарид с конкретными физическими свойствами (температура плавления, растворимость и т.д.) характеризуется специфической величиной удельного вращения [о|о20.
О./
тлш
I
с^о
I
СН|ОН
Дшешцвтш
I
гн .он
1 *
с -о неон
I
снаон
1>шшшк»за ^r ’ -
с—о
1
неон
i
нош
J
неон
1
СН2ОН
0«Ш|Лм
«гон
1
сн,он
D-псщом
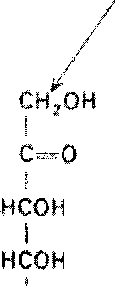


СН'аОН
I
с—о
неон
(
тт
|
сн2ом
0‘рй%яоаа
Ш2ОН I С-0
НОСН
I
ИСОН
!
неон
1
снгон
©«фруктоза
скон 1 '
носн
I
штп
1
СН2ОК
С—О
I
НОСН
I
носн
I
неон
I
CHjOH
0«тагатоза
Явление мутаротации имеет объяснение. Известно, что альдегиды и ке-тоны легко и обратимо реагируют с эквимолярным количеством спирта с образованием полуацеталей:
I
fteipitiiii
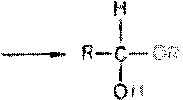
Реакция образования полуацеталя возможна и в пределах одной молекулы, если это не связано с пространственными ограничениями. По теории А. Байера, внутримолекулярное взаимодействие спиртовой и карбонильной групп наиболее благоприятно, если оно приводит к образованию пяти- или шестичленных циклов. При образовании полуацеталей возникает новый асимметрический центр (для D-глюкозы это С-1). Шестичленные кольца сахаров называют пиранозами, а пятичленные - фуранозами. а-Форма - это форма, у которой расположение полуацетального гидроксила такое же, как у асимметричного углеродного атома, определяющего принадлежность к D- или L-ряду. Иными словами, в формулах с а-моди-фикацией моносахаридов D-ряда полуацетальный гидроксил пишут справа, а в формулах представителей L-ряда - слева. При написании |3-формы поступают наоборот.
Таким образом, явление мутаротации связано с тем, что каждый твердый препарат углеводов представляет собой какую-либо одну циклическую (полуацетальную) форму, но при растворении и стоянии растворов эта форма через альдегидную превращается в другие таутомерные циклические формы до достижения состояния равновесия. При этом значение удельного вращения, характерное для исходной циклической формы, постепенно меняется. Наконец, устанавливается постоянное удельное вращение, которое характерно для равновесной смеси таутомеров. Например, известно, что в водных растворах глюкоза находится главным образом в виде а- и (3-глюкопираноз, в меньшей степени - в виде а- и (3-глюкофураноз и совсем небольшое количество глюкозы - в виде альдегидной формы.
Следует подчеркнуть, что из различных таутомерных форм глюкозы в свободном состоянии известны лишь а- и p-пиранозы. Существование малых количеств фураноз и альдегидной формы в растворах доказано, но в свободном состоянии они не могут быть выделены вследствие своей неустойчивости.
Установлено, что величина удельного вращения при растворении любого моносахарида постепенно меняется и лишь при дли тельном стоянии раствора достигает вполне определенного значения. Например, для свежеприготовленного раствора глюкозы [а] о = +112,2°, после длительного стояния раствора эта величина достигает равновесного значения [а]о20= +52,5°. Изменение величины удельного вращения при стоянии (во времени) растворов моносахаридов называется мутаротацией. Очевидно, мутаротация должна вызываться изменением асимметрии молекулы, а следовательно, трансформацией ее структуры в растворе.
В 20-х годах У. Хеуорс предложил более совершенный способ написания структурных формул углеводов. Формулы Хеуорса - шести- или пятиугольники, причем они изображены в перспективе: кольцо лежит в горизонтальной плоскости. Находящиеся ближе к читателю связи изображают более жирными линиями (углеродные атомы цикла не пишут). Заместители, расположенные справа от остова молекулы при ее вертикальном изображении, помещают ниже плоскости кольца, а заместители, находящиеся слева- выше плоскости кольца. Обратное правило применяют только для того единственного углеродного атома, гидроксильная группа которого участвует в образовании
циклического полуацеталя. Так, у D-сахаров группу СНгОН пишут над этим атомом углерода, а водородный атом при нем - внизу.
Наконец, следует помнить, что при написании структурных формул по Хеуорсу гидроксильная группа при С-1 должна быть расположена ниже плоскости кольца в а- форме и выше - в (3-форме:
в-!3-гш«пирамоа*
^-0-гл«ясй-ира1нрэ8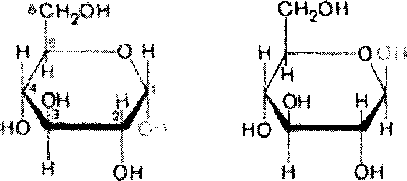
щ
I
«эм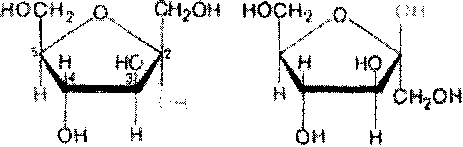
![]()
Проекционные формулы Хеуорса не отражают подлинной конформации моносахаридов. Подобно циклогексану, пиранозное кольцо может принимать две конфигурации - форму кресла и форму лодки (конформацион-ные формулы). Форма кресла обычно более устойчива, и, по-видимому, именно она преобладает в большей части природных сахаров
ОЛИГОСАХАРИДЫ
Олигосахариды - углеводы, молекулы которых содержат от 2 до 10 остатков моносахаридов, соединенных гликозидными связями. В соответствии с этим различают дисахариды, трисахариды и т.д.
Дисахариды - сложные сахара, каждая молекула которых при гидролизе распадается на две молекулы моносахаридов. Дисахариды наряду с полисахаридами являются одними из основных источников углеводов в пище человека и животных. По строению дисахариды - это гликозиды, в которых 2 молекулы моносахаридов соединены гликозидной связью.
ц
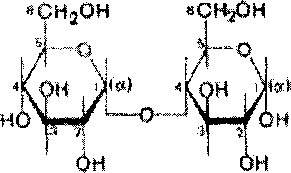
Мшоав
В молекуле мальтозы у второго остатка глюкозы имеется свободный полуацетальный гидроксил. Такие дисахариды обладают восстанавливающими свойствами.
Одним из наиболее распространенных дисахаридов является сахароза - обычный пищевой сахар. Молекула сахарозы состоит из одного остатка D-глюкозы и одного остатка D- фруктозы. Следовательно, это а-глюко-пиранозил-(1->2)-|3-фруктофуранозид:
ОМ|СШ

В отличие от большинства дисахаридов сахароза не имеет свободного полуацетального гидроксила и не обладает восстанавливающими свойствами. Гидролиз сахарозы приводит к образованию смеси, которую называют инвертированным сахаром. В этой смеси преобладает сильно левовращающая фруктоза, которая инвертирует (меняет на обратный) знак вращения правовращающего раствора исходной сахарозы.
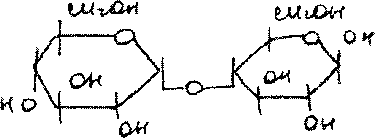
Дисахарид лактоза содержится только в молоке и состоит из D-галак-тозы и D-глюкозы. Это - Р-галактопиранозил-(1->4)-глюкопираноза:
9К4ОН ОДОЙ '
Ames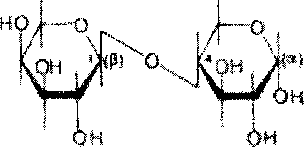
Благодаря наличию в молекуле свободного полуацетального гидрокси-ла (в остатке глюкозы) лактоза относится к числу редуцирующих ди-сахаридов.
Среди природных трисахаридов наиболее известна рафиноза, содержащая остатки фруктозы, глюкозы и галактозы. Рафиноза в больших количествах содержится в сахарной свекле и во многих других растениях. В целом олигосахариды, присутствующие в растительных тканях, разнообразнее по своему составу, чем олигосахариды животных тканей.
Полисахариды. Г омополисахариды.
По своему функциональному назначению гомополисахариды могут быть разделены на две группы: структурные и резервные полисахариды. Важным структурным гомополисахаридом является целлюлоза, а главными резервными - гликоген и крахмал (у животных и растений соответственно).
Г^ТТЛПГЯСТ ТГ ТТЯРП/ГгЬиТГЯТТТ/Га Т7Г» YITA/fT/HJPPVnA/TV Т/ТГГЪГ ГЛТ/ТП ТТГ\Т"ТТи(^Г‘ТГГ\Т^ 1ЛГЧТТТТ 1ЭГ» ТТ#=» ТТГ»гГОТЖ/=»
1V.J ч», 1J.V /1I11UI1 ivvnvmj VlpV/ViilllV 11^111 \J XXV/ J1V71 XX ~X WV1W XX IDIl V
отсутствия для многих полисахаридов исчерпывающих данных невозможна. Поэтому чаще всего полисахариды «именуются» по источникам выделения, несмотря на то что один и тот же полисахарид может быть получен из совершенно разных источников.
Крахмал, как отмечалось, является основным резервным материалом растительных организмов. В небольших количествах он содержится в листьях, но главным образом накапливается в семенах (зерна злаков, например пшеницы, риса, кукурузы, содержат до 70% крахмала), а также в луковицах, клубнях и сердцевине стебля растений, где содержание его доходит до 30%.
Крахмал представляет собой смесь 2 гомополисахаридов: линейного - амилозы и разветвленного - амилопектина, общая формула которых (СбНю05)п. Как правило, содержание амилозы в крахмале составляет 10-30%, амилопектина - 70-90%. Полисахариды крахмала построены из остатков D-глюкозы, соединенных в амилозе и линейных цепях амило-пектина а-1->4-связями, а в точках ветвления амилопектина - межцепочечными а-1->6-связями:
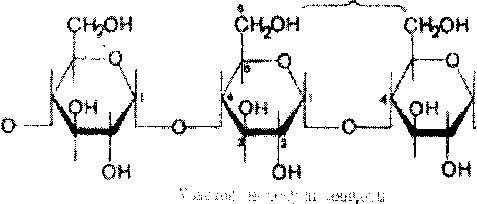
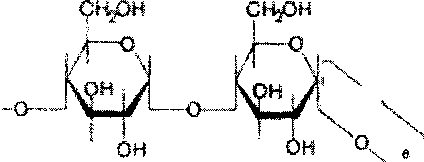
о
\
I \|
н<М,*Ом»
. ш
Ш
I(
он
#нусмн
—о
![]()




Итак, единственным моносахаридом, входящим в состав крахмала, является D-глюкоза. В молекуле амилозы линейно связано в среднем около 1 ООО остатков глюкозы; отдельные участки молекулы амилопектина состоят из 20-30 таких единиц. В настоящее время общепринятой является «ветвистая» структура отдельных цепочек с а-1->4-связями в молекуле амилопектина (рис. 5.3).
Известно, что в воде амилоза не дает истинного раствора. Цепочка амилозы в воде образует гидратированные мицеллы. В растворе при добавлении йода амилоза окрашивается в синий цвет. Амилопектин также дает мицеллярный раствор, но форма мицелл несколько иная. Полисахарид амилопектин окрашивается йодом в краснофиолетовый цвет.
§ 9 \ё *
HQ€t%
HtOGI% NeHjf HOCtfe
:0N*“
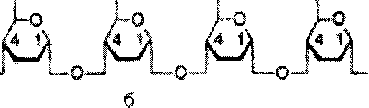

![]()
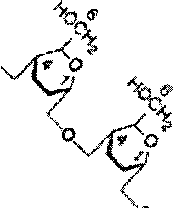
Рис.
5.3. Структура крахмала: а - амилоза с
характерной для нее спиральной
структурой; б - амилопектин, образующий
в точках ветвления связи типа 1-6.
&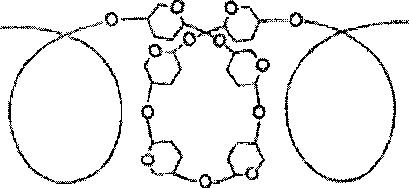
Рис.
5.4. Строение
отдельного участка (а) и всей молекулы
(б) гликогена (по Майеру). Белые кружки
- остатки глюкозы, соединенные
а-1,4-связью; черные кружки - остатки
глюкозы, присоединенные а-1,6-связью; R
- редуцирующая концевая группа. Внутренние
цепи, или ветви,- участки между точками
ветвления. Наружные цепи, или ветви,
начинаются от точки ветвления и кончаются
нередуцирующим остатком глюкозы.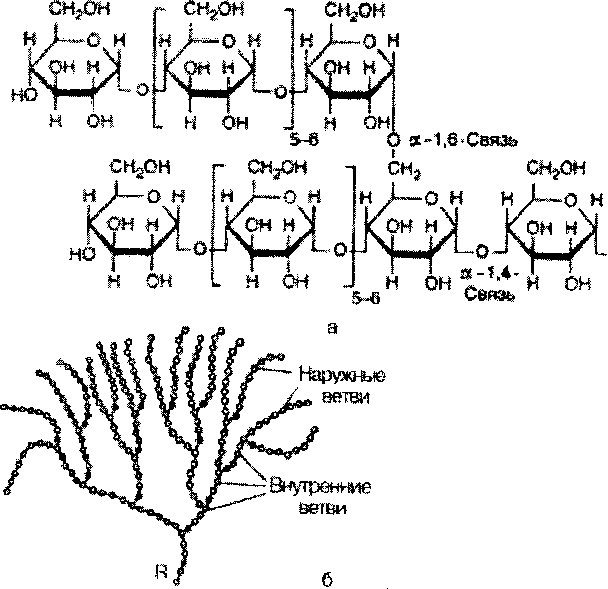
Для человека крахмал является важным пищевым углеводом; содержание его в муке составляет 75-80%, в картофеле - 25%. '
Гликоген - главный резервный полисахарид высших животных и человека, построенный из остатков D-глкжозы. Эмпирическая формула гликогена, как и крахмала, (C6Hi0O5)n. Гликоген содержится практически во всех органах и тканях животных и человека; наибольшее количество обнаружено в печени и мышцах. Молекулярная масса гликогена 10-10 Да и более. Его молекула построена из ветвящихся полиглюкозидных цепей, в которых остатки глюкозы соединены а-1->4-гликозидными связями. В точках ветвления имеются а-1 ->6-гликозидные связи. По строению гликоген близок к амилопектину. В молекуле гликогена различают внутренние ветви - участки от периферической точки ветвления до нередуци-рующего конца цепи (рис. 5.4).
Гликоген характеризуется более разветвленной структурой, чем амило-пектин; линейные отрезки в молекуле гликогена включают 11-18 остатков a-D-глюкопиранозы.
При гидролизе гликоген, подобно крахмалу, расщепляется с образованием сначала декстринов, затем мальтозы и, наконец, глюкозы.
t
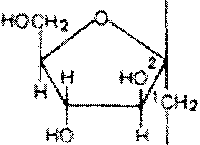
Метилирование инулина свидетельствует, что остатки D-фруктозы связаны между собой
>1-связями и находятся в фуранозной форме:
Шшггж
дымирш»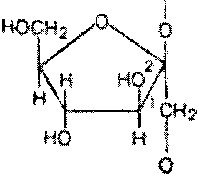

Хитин - важный структурный полисахарид беспозвоночных животных (главным образом членистоногих). Из него, в частности, построен наружный скелет ракообразных и насекомых.
Хитин также частично или полностью замещает целлюлозу в клеточных стенках сапрофитных растений, например грибов.
Структуру хитина составляют Ы-ацетил-О-глкжозаминовые звенья, соединенные р-(1- >4)-гликозидными связями:
Повкуявщй«:§
звенья в жшщяш
ша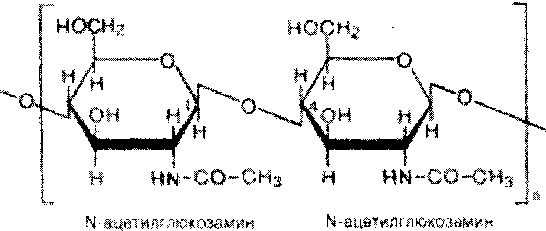
Целлюлоза (клетчатка) - наиболее широко распространенный структурный полисахарид растительного мира. Он состоит из а-глюкозных остатков в их Р-пиранозной форме, т.е. в молекуле целлюлозы Р-глюко-пиранозные мономерные единицы линейно соединены между собой Р-(1->4)-связями:
![]()
щрн снрн
I I ж-
—о
1 Off 1 6h
J«
Лжяш mimym mmm&rn
При частичном гидролизе целлюлозы образуется дисахарид целлобиоза, а при полном гидролизе D-глюкоза. Молекулярная масса целлюлозы 1000-2000 кДа. Клетчатка не переваривается ферментами пищеварительного тракта, так как набор этих ферментов v человека не содержит гидролаз, расщепляющих p-связи. В связи с этим целлюлозу можно рассматривать как значительный неиспользуемый «пищевой» резерв. Вместе с тем известно, что присутствие оптимальных количеств клетчатки в пище способствует формированию кала. При полном исключении клетчатки из пищи нарушается формирование каловых масс.
В кишечнике жвачных и других травоядных животных имеются микроорганизмы способные к ферментативному расщеплению P-связей (р-глю-козидных связей) и для этих животных целлюлоза является важным источником пищевых калорий. ’
Наконец, целлюлоза и ее производные имеют колоссальное практическое значение. Основная масса целлюлозы используется для изготовления хлопчатобумажных тканей и бумаги. Кроме того, на основе целлюлозы производятся искусственные волокна пластмассы и т.д. Характерной особенностью целлюлозы, определяющей в значительной степени ее механические, физико-химические и химические свойства, является линейная конформация молекул, закрепленная внутримолекулярными водородными связями.
Г етерополисахариды
Полисахариды, в структуре которых характерно наличие двух или более типов мономерных звеньев, носят название гетерополисахаридов.
Принято считать, что, поскольку гетерополисахариды чаще состоят только из двух различных мономеров, расположенных повторяющимся образом, они не являются информационными молекулами [Бохински Р., 1987].
Важнейшие представители гетерополисахаридов в органах и тканях животных и человека гликозаминогликаны (мукополисахариды). Они состоят из цепей сложных углеводов содержащих аминосахара и уроновые кислоты. ’
р-Глэ»сршовая
«сшта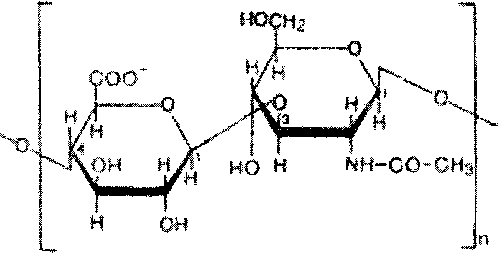
Хондрс»мтш***<срь«|»т
|eeip®4ae?cs
т«жв#*ерьфй}
|?-Гя1!М£рюж®ая М-*1Р:ти11г»«т«-
«Wafa амиигрьфйг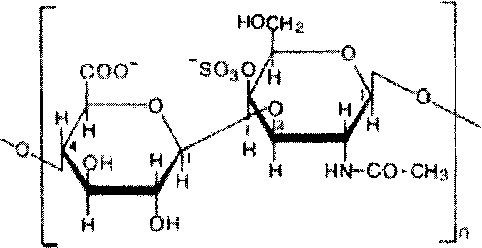
Гешрин
№0080** И
гштаашн «дуренша» кмешга
Рис.
5.5.
Строение
некоторых сложных полисахаридов
(гликозамино-гликанов).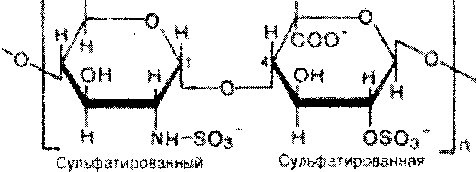
![]()
Рис.
5.6. Протеогликановый
агрегат (схема).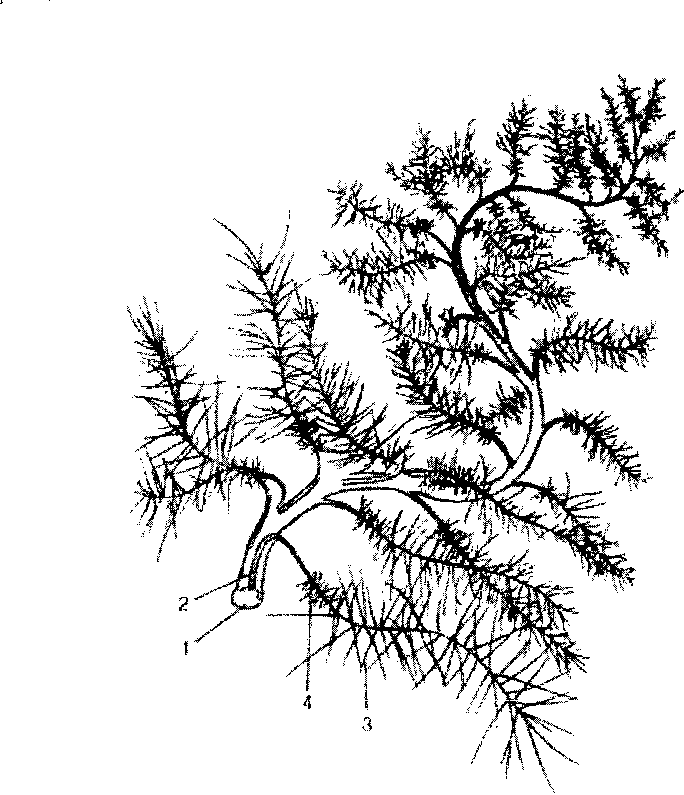
Единая длинная молекула гиалуро-ната (1) нековалентно связана со многими молекулами белка (2), каждая из которых содержит кова-лентно связанные молекулы хонд- роитинсульфата (3) и кератансуль-фата (4).
Различают шесть основных классов гликозаминогликанов .Каждый из гликозаминогликанов содержит характерную для него повторяющуюся дисахаридную единицу; во всех случаях (кроме кератансульфатов) эта единица содержит либо глюкуроновую, либо идуроновую кислоту. Все гликозаминогликаны, за исключением гиалуроновой кислоты, содержат остатки моносахаридов с О- или N-сульфатной группой.
Гликозаминогликаны значительно различаются по размерам, их молекулярные массы в пределах от 104 Да для гепарина до 107 Да для гиалуроновой кислоты.
Выделенные индивидуальные гликозаминогликаны могут содержать смесь цепей различной длины (рис. 5.5). Гликозаминогликаны как основное скрепляющее вещество связаны со структурными компонентами костей и соединительной ткани. Их функция состоит также в удержании большой массы воды и в заполнении межклеточного пространства. Иными словами, гликозаминогликаны — основной компонент внеклеточного вещества — жела-тинообразного вещества, заполняющего межклеточное пространство тканей. Они также содержатся в больших количествах в синовиальной жидкости - это вязкий материал, окружающий суставы, который служит смазкой и амортизатором.
Поскольку водные растворы гликозаминогликанов гелеобразны, их называют мукополисахаридами.
Наконец, если цепи гликозаминогликана присоединены к белковой молекуле, соответствующее соединение называют протеогликаном.
Протеогликаны образуют основное вещество внеклеточного матрикса. В отличие от простых гликопротеинов, которые содержат только несколько процентов углеводов (по массе), протеогликаны могут содержать до 95% (и более) углеводов (рис. 5.6).
Обмен и функции углеводов (лекционный материал)
Cn(H20)m
Углеводы (УВ)- альдегиды и кетоны многоатомных спиртов и их полимеры. На долю углеводов приходится 2% от массы тела.
Функции:
Энергетическая -50-60% Е за счёт УВ; При полном окислении 1 г УВ обр-ся 4.1 ккал Е
Пластическая -из УВ в организме синтезируются белки,н.к,липиды.
Защитная -глюкозоаминогликан-гиалур.к-та У находится в соединительной ткани,образуют сетеобразные структуры, препятствующие проникновению чужеродныж в-в. Покрывают эпителий дых.путей,обр.гликокаликс кишечника,роль смазочного материала в суставах.
Опорная,механическая- в составе соед.ткани участв-ет в построении опорно - двигательного аппарата.
Гидраосмотическая,ионрегулирующая- в составе УВ много OH-rpynn,V притягивают воду,удерживают её.Притягивают катионы.
Поддержание гемостаза-гепарин участвует в антисвертывающей системе крови.
Резервное депонирование-откладывается про запас -гликоген-энерг.пластичный материал. Обнаружен во всех клетках,больше всего в печени и мышцах.У жен. лактоза грудного молока.
Обезвреживающая функция-глюкуроновая кислота участвует в процессе обезвреживания токсичных в-в(своих)и ксенобиотиков(из внешней среды).
Специфическая функция -Радикальная ф-ия- R содержат УВ(гликопротеиды)
Регуляторная -гормоны гипофиза(гликопротеиды).
Защитная -при формировании иммунитета(В форме антител).
Потребность:
Для взрослого человека 400-600г(5,5-7г/кг)
У ребёнка потребность больше 10-15г/кг 50-100г УВ:
-продукты растительного происх-я -крахмал(хлеб,крупы и др)
-в животных продуктах-гликоген(мясо,печень,почки)
-дисахариды-в молоке -сахароза-сахар,ананас.морковь -мальтоза-проросшие семена злаков,пиво,солод.
Клетчатка-в оболочках кл. растений,не гидролизуется вЖКТ.
Значения клетчатки:
![]()
Удерживает воду в кишечнике,формируя мягк.фекалии,что очень важно для старых людей.
тормозит всасывание холестерина из кишечника,уменьшает его уровень в крови,тормозит развитие атеросклероза
обеспечивает медл.поступление глюкозы в кровь,тормозится секреция инсулина.
Обмен ув
Идет в 3 этапа:
)Подготовительный-пищеварение(поступление—^расщепление в ЖКТ—>всас-е)
Внутриклеточный обмен(Катаболизм,Анаболизм)
Образование конечных продуктов,выведение конечных продуктов
Превращене УВ осущ-я / 3 класса гидролаз-1 ликозидаз: 1 )эндогл икозидазы,^ гидролизуют внутр.гликозидные связи,2)экзогликозидазы,^ расщ.наружные гликозидные связи. В ходе переваривания поли-олиго-дисахариды расщепляются до моносахаридов,V всасываются.
Превращение полисахаридов.
В ротовой полости на полисахариды действует слюна(95.5% воды ,/,белки),в её составе / q-амилаза. По своему действию-эндогликозидазы гидролизуют а1,4-гликозидные связи,рН=6,8-7,2. Полисахариды ращепляются до декстринов,V всасываться не могут и поступают в желудок. В составе слюны есть /-мальтаза. В желудке рН=1,5-2(кислая),НС1 ингибирует а-амилазу слюны и переваривание полисахаридов может продолжаться внутри пищевого комка под действием а-амилазы,до тех пор, пока HCI не пропитает весь комок, /-ов,переваривающих УВ в желудке -нет!=> 12кишке рн=7,5-8(щел),основной / - а-амилаза пакреотического сока,эндоамилаза расщепляет внутренние а-1,4 глик.связи. Кроме этих / в 12 кишке дейс-ет /, V расщепляет а 1,6-связи: амило-1,6- гликозидаза,олиго-1,6-гликозидаза-/ кишечного сока. Амило-1,6-глюкозидаза гидролизует декстрины,содержит более 10-12 остатков глюкозы. Олиго-1,6-глю-за-до 1012 ост.глюкозы. Итогом работы всех вышеуказанных / будет образование из полисахаридов дисахариды-мальтаза,изомальтаза—^переваривание дисахаридов в кишечнике.
tUvOH
НО
CWtOH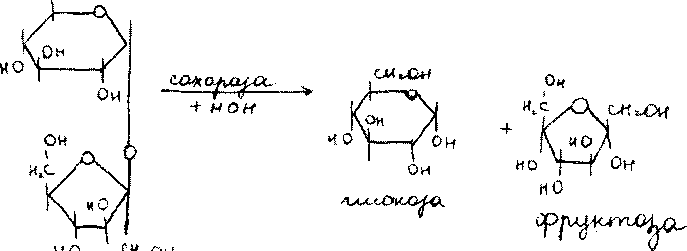
о
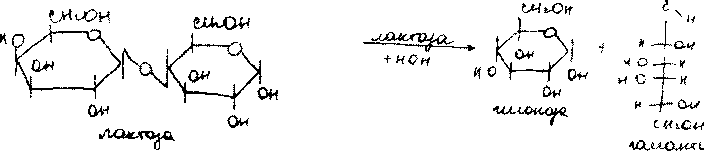
«ш
 4
и*”""
4
и*”""
-4 «ои ™
цо
ОН ЩАЛЛШШ^Я
Переваривание дисахаридов идёт на пов-и энтероцита(пристеночное пищ-е) / работают не изолированно^ образуют комплексы:
Сахаразнаизомальтазный
Р-гликозидазный
Гликоамилазный(/-мальтаза) ,
/А, ' 1: -ДютмМШМ
'V . :
f, - \ .„I: 1*"? '
Особенности переваривания УВ у детей.
Основной УВ-лактоза(молоко)По мере роста и развития добавл. гликоген,крахмал. /-а- амилаза слюны и подж.железы обладают меньшей активностью, а-амилаза слюны достигает нормы к 2-7 годам ,а подж.жел. к 3-5г. Основной /^переваривает дисахариды- лактаза,наиболее активна,активность привышает активность у взрослых. При переваривании полисахаридов в желудке у детей среда менее кислая 3,5-4,5,гидролиз полисахаридов глубокий ,полный.
Всасывание может идти:
путём пассивной диффузии по градиенту концентрации-галак. и глю при голодании
При |конц.глю в крови она идёт против градиента конц, -активный транспорт. Участвуют АТФ,К-№-АТФаза,натрий. У детей проницаемость кишечной стенки выше,чем у взрослого. Пост, и моно и дисахариды,декстрины. Особенностью работы киш.детей до 1 года-брожения УВ. В результате брожения образуются органические кислоты,V стимулируют перистальтику киш-ка,способствуют всасыванию продуктов расщепления УВ. Основ.микролатобактерии-lactobacillus bifidus. Бифидобактерии участвуют в синтезе нек-х витаминов(К,В1),подавляют процессы гниения. Для роста бактерий необходим bifidus-фактор-совокупность в-в,обеспеч-х рост бактсрий(УЮ сод-я в жен.молоке)глю,(3-латоза-дольше сохраняется в кишечнике-пища для бактерий))
Глюкоза в крови.
Более пловины Глю(60%)всасывается по портальной вене—^печень,где 2\3 превращается в гликоген,а 1\3-в жиры,окисляется. 40% Глю с кровотока поступает в мышцы и жир.тк.
В мышце происходит синтез гликогена,в жир.тк-синтез жира. Часть Глю из общего русла—>мозг,эритроциты(инсулин независимые ткани) Поступление глю в кл. с помощью белков-переносчиков ГЛЮТ:
ГЛЮТ 1 -переносит глю в мозг,эритроциты ГЛЮТ 2-в печень,кл.кишечника ГЛЮТ 3-в нервн.тк.
ГЛЮТ 4 -мышцы,жир.тк ГЛЮТ 5-тонк.к-к
В норме конц-я Глю в крови -60-100мл\дл,3,33-5,55 мммоль\л(3,5-6,1 ммоль\л) Пути поступления и испльзования глю крови.
УВ пиар
I
Распад
гликогена в печени
Глюкоза
крови
Глюконеогенез{гнг)
Синтез
гликогена в
гликогена
в печени
дихотомическое
2ПВК *02}
2
лактата 2АТФ
С02,38(36-38)
ДТФ
Н20








Виды и пшчины гипоГгипеп^гликемии
• л. \ Г/
Гипергликемия- увеличение конц-ии Глю в крови выше 5,55 ммоль\л
Физиологическая -алиментарная
-стрессорная (сихоэмоциональная)
Патологическая
-усиленная выработка глик.гормонов(феохромацитон н\п,опухоль а клеток подж.жел,тереотоксидоз,при сахарном диабете)
Гипогликемия- снижение конц.глю в крови ниже 3,33 ммоль\л
Физиологическая -интенсивная мыш.работа -беременность -лактация -голодание
-у астеников
-неонатальная(у ребёнка сразу после рождения ч\з 4-6 часов после родов,глю падает до 2,2 ммоль\л и восстан.до нормы к 10-14 годам
Патологическая -опухоли кл Р подж.ж(инсулома)
-гликогенозы -пор-я ЖКТ
При | конц.глю в крови выше 8,8 ммоль\л она выделяется с мочой. Эта концентрация глю наз. почечным пороком .(тубулярный мах)
Глюкостатическая ф-ия печени.
Важную роль в поддержании постоянного уровня глю крови отводится печени-ф-ия глюкостатическая. Эта ф-я осущ-я 2 причинами:
При уменьшении конц глю в крови —>увел-тся распад гликогена в печени.
При повышении глю в крови —^усиливается синтез гликогена в печени.
Синтез гликогена (гликогеногенез).
Общее кол-во гликогена 5 ООг,больше всего в мышцах,в печени -6% от массы Процесс синтеза начинается ч\з 1-2 часа после приёма пищи,связан с затратой Е.
cUv~oH
.
„ » ^ Ли AIT I \t SS о« ....
40
т f | f 1 ОИ
ш*о
'LU/WJ 4juuX>-
t>~
V ио
Mf с,—_
Л
'Л, —■*
он
, И К Cm л
ШьОИ f +
![]()






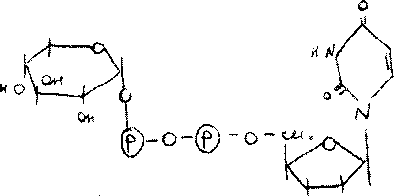
/-глюкокиназа переводит глю в фосфорилированную форму и запирает глю в клетке. Свободная глю легко входит в кл и выходит ,а фосф.эфир из кл.выйти не может,не ингиб. глю-6-фосфатом.
Синтез этого / актив.гармон инсулин
(йн^Об), * ticfw.
Гликоген затравка-то мин.кол-во гликогена,V всегда остаётся в гепатоцитах,как трудно организму не пришлось,в основе -Б-гликогенин,к^ присое-я не менее 8 остатков глю. Реакция^ катализ, гликогенсинтаза-р-я необратимая. Гликогенсинтаза - ключевой / синтеза гликогена.
&
С4
С1
V
—и
Тостатков
глю
/ ветвления начинает действовать с нередукционного конца,отщепл.7остатков глю и переносит к середине оставшиеся цепи,образответвления,доп-но а-1,6 связь дальше синтез
гликогена продолжается.

ооооо
Значение:
Молекула становится более компактной
Увеличивается кол-во нередуц-х концов(все / работ, с нередуц.концов)
Молекула гликогена становится более гидрофильной
в составе развлет-ой молекулы запасается большое кол-во глю.
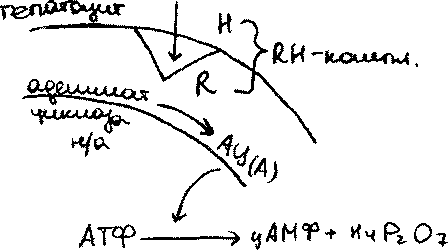
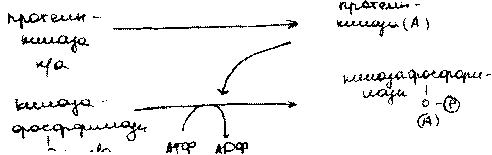
Распад гликогена в печени (мобилизация гликогена в печени, гликогенолиз).
а процессе распада гликогена в печени образуется свободная rjuo.V поступает в кровь и поддерживает постоянный уровень глю в крови,Сигналом для распада гликогена служит пониж.концГлю в крови > выброс гармона глюкогона,V запускает процессы активации / гликогенфосфорилаза. Пр. распада гликогена может быть вызван адреналином(при
Схема активации ГФ-зы. Распад гликогена в печени.
А У
Тостатков глю
ВСТВЯеНИСТ НЯХПТНЯР'Г ТТР‘Т^Г‘гггзглпо'г-т
' ~ ^vxxviuv.uuid v псред^кциилниго конца,отщепл. /остатков глю и
переносит к середине оставшиеся цепи,образответвления,доп-но а-1,6 связь дальше синтез гликогена продолжается.
С4 г»-»
ооооо
Значение:
Молекула становится более компактной
Увеличивается кол-во нередуц-х концов(все / работ, с нередуц.концов)
Молекула гликогена становится более гидрофильной
в составе развлет-ой молекулы запасается большое кол-во глю.
Распад гликогена в печени (мобилизация гликогена в печени, гликогенолиз).
а процессе распада гликогена в печени образуется свободная глюЛ поступает в кровь и поддерживает постоянный уровень глю в крови,Сигналом для распада гликогена служит пониж.концГлю в крови >выброс гармона глюкогона,V запускает процессы активации / гликогенфосфорилаза. Пр. распада гликогена может быть вызван адреналином(при слрсссс)
г

т игы ,
%
гшАягеллм ■
f» Vх» 3
(йн^Об), * ticfw. 22
<т®<®т<8соосо 23
ооооо 23
ооооо 25
амщ 27
к... -Дг, АТЧР АРФ
АДФ
Н20 тишзгенсшгаза
I н\а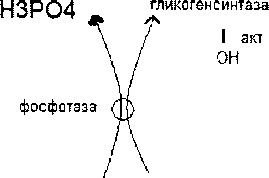
«ваза
АТФ
![]()