

12)Классификация рецепторов по способу передачи сигнала. G-белок ассоциированный рецептор, принципы передачи внутриклеточного сигнала, примеры фармакологических лигандов.
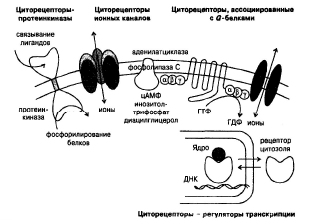
Циторецепторы классифицируют на 4 типа (рис. 3):
· рецепторы-протеинкиназы;
· рецепторы ионных каналов;
· рецепторы, ассоциированные с G-белками;
· рецепторы-регуляторы транскрипции.
Циторецепторы, ассоциированные с G-белками, представляют собой группу интегральных
мембранных белков с характерной вторичной структурой, включающей внеклеточный N-конец и
внутриклеточный С-конец, 7 трансмембранных доменов, связанных тремя внеклеточными и тремя
внутриклеточными петлями. Белковая цепь циторецепторов содержит большое количество остатков
цистеина, образующих дисульфидные мостики. Внеклеточные петли, выполняя регуляторную
функцию, ответственны за распознавание и связывание лигандов. G-белки, расположенные внутри
мембраны, передают информацию от внеклеточного регуляторного домена на эффекторную систему,
используя энергию ГТФ. Внеклеточные и трансмембранные домены согласованно участвуют в
связывании лигандов и активации циторецепторов.
Эффекторная система представлена аденилатциклазой, фосфолипазами А2, С и D, белками ионных
каналов, транспортными белками. При возбуждении рецепторов образуются внутриклеточные
биологически активные вещества — вторичные мессенджеры (англ. messenger — вестник,
посредник).
Аденилатциклаза превращает АТФ во вторичный мессенджер цАМФ. Рецепторы могут как
активировать (RS), так и ингибировать (Ri) аденилатциклазу.
13)Классификация рецепторов по способу передачи сигнала. Рецепторные протеинкиназы, принципы передачи внутриклеточного сигнала, примеры фармакологических лигандов.
рецепторы-регуляторы транскрипции.
Циторецепторы классифицируют на 4 типа (рис. 3):
• рецепторы-протеинкиназы;
• рецепторы ионных каналов;
• рецепторы, ассоциированные с G-белками;
• рецепторы-регуляторы транскрипции.
Циторецепторы-протеинкиназы связаны с плазматической мембраной клеток. Они имеют
внеклеточный домен для взаимодействия с лигандами и внутриклеточный каталитический домен —
протеинкиназу. Оба домена соединены внутримембранной цепью из гидрофобных аминокислот.
Рецепторы-протеинкиназы фосфорилируют белки клеток — киназы, регуляторные и структурные
белки. Чаще всего происходит фосфорилирование тирозина белков-мишеней. Примеры
циторецепторов-протеинкиназ — рецепторы инсулина, цитокинов, эпидермального и
тромбоцитарного факторов роста.
Циторецепторы к предсердному (атриальному) натрийуретическому пептиду, ассоциированные с
гуанилатциклазой, повышают продукцию цГМФ.
14)Классификация рецепторов по способу передачи сигнала. Ионотропые рецепторы, принципы передачи внутриклеточного сигнала, примеры фармакологических лигандов.
Циторецепторы ионных каналов, повышая проницаемость мембран для Na+, K+, Са2+ и Сl-,
обеспечивают мгновенный клеточный ответ. Примеры рецепторов ионных каналов:
· рецепторы, вызывающие деполяризацию и возбуждение функций клеток, — Н-холинорецепторы,
рецепторы глутаминовой и аспарагиновой кислот (увеличивают проводимость для Na+, K+ и Са2+);
· рецепторы, вызывающие гиперполяризацию и торможение функций клеток, — ГАМКА-рецепторы
и рецепторы глицина (увеличивают проводимость для Сl-).
15))Классификация рецепторов по способу передачи сигнала. Внутриклеточные рецепторы, регулирующие транскрипцию генов. Принципы передачи внутриклеточного сигнала, примеры фармакологических лигандов.
Циторецепторы классифицируют на 4 типа (рис. 3):
• рецепторы-протеинкиназы;
• рецепторы ионных каналов;
• рецепторы, ассоциированные с G-белками;
• рецепторы-регуляторы транскрипции.
Циторецепторы-регуляторы транскрипции взаимодействуют с тиреоидными, стероидными
гормонами, витамином D и ретиноидами. Транспортные белки крови передают лиганды клетоным
белкам, затем комплексы поступают в ядро. Функции рецепторов — активация или ингибировние
транскрипции генов.Рецепторную функцию выполняют также мембраносвязанные и растворимые ферменты(дигидрофолатредуктаза, ацетилхолинэстераза, моноаминоксидаза, циклоксигеназа), транспортные белки (Na+, К+-АТФ-аза) и структурные белки (тубулин).
16)Типы, структура и локализация M-холинорецепторов. Передача внутриклеточного сигнала и эффекты, сопряженные с активацией М-холинорецепторов.
Холинергические синапсы локализованы в ЦНС (ацетилхолин регулирует моторику,
пробуждение, память, обучение), а также в вегетативных ганглиях, мозговом слое надпочечников,
каротидных клубочках, скелетных мышцах и внутренних органах, получающих постганглионарные
парасимпатические волокна
Молекулярное клонирование позволило выделить пять типов М-холинорецепторов:
1. М1-холинорецепторы ЦНС (лимбическая система, базальные ганглии, ретикулярная
формация) и вегетативных ганглиев;
2. М2-холинорецепторы сердца (снижают частоту сердечных сокращений, атриовентрикулярную
проводимость и потребность миокарда в кислороде, ослабляют сокращения предсердий);
3. М3-холинорецепторы:
· гладких мышц (вызывают сужение зрачков, спазм аккомодации, бронхоспазм, спазм
желчевыводящих путей, мочеточников, сокращение мочевого пузыря, матки, усиливают
перистальтику кишечника, расслабляют сфинктеры);
· желез (вызывают слезотечение, потоотделение, обильное отделение жидкой, бедной белком
слюны, бронхорею, секрецию кислого желудочного сока).
Внесинаптические М3-холинорецепторы находятся в эндотелии сосудов и регулируют
образование сосудорасширяющего фактора — окиси азота (NО).
4. М4- и М5-холинорецепторы имеют меньшее функциональное значение.
М1-, М3- и М5-холинорецепторы, активируя посредством Gq/11-белка фосфолипазу С клеточной
мембраны, увеличивают синтез вторичных мессенджеров — диацилглицерола и
инозитолтрифосфата. Диацилглицерол активирует протеинкиназу С, инозитолтрифосфат
освобождает ионы кальция из эндоплазматического ретикулума,
М2- и М4-холинорецепторы при участии Gi- и G0-белков ингибируют аденилатциклазу (тормозят
синтез цАМФ), блокируют кальциевые каналы, а также повышают проводимость калиевых каналов
синусного узла.
17)Типы, структура и локализация N-холинорецепторов. Передача внутриклеточного сигнала и эффекты, сопряженные с активацией N-холинорецепторов.
Н-холинорецепторы широко представлены в организме. Их классифицируют на Н-
холинорецепторы нейронального (Нн) и мышечного (Нм) типов.
Нейрональные Нн-холинорецепторы представляют собой пентамеры и состоят из субъединиц a2 —
a9, и β2 — β4 (четыре трансмембранные петли). Локализация нейрональных Н-холинорецепторов
следующая:
· кора больших полушарий, продолговатый мозг, клетки Реншоу спинного мозга, нейрогипофиз
(повышают секрецию вазопрессина);
· вегетативные ганглии (участвуют в проведении импульсов с преганглионарных волокон на
постганглионарные);
· мозговой слой надпочечников (повышают секрецию адреналина и норадреналина);
· каротидные клубочки (участвуют в рефлекторном тонизировании дыхательного центра).
Мышечные Нм-холинорецепторы вызывают сокращение скелетных мышц. Они представляют
собой смесь мономера и димера. Мономер состоит из пяти субъединиц (a1 — a2, β, γ, ε, δ),
окружающих ионные каналы. Для открытия ионных каналов необходимо связывание ацетилхолина с
двумя a-субъединицами.
Н-холинорецепторы возбуждаются алкалоидом табака никотином в малых дозах, блокируются
никотином в больших дозах.
Биохимическая идентификация и выделение Н-холинорецепторов стали возможны благодаря
открытию их избирательного высокомолекулярного лиганда a-бунгаротоксина — яда тайваньской
гадюки Bungarus multicintus и кобры Naja naja. Н-холинорецепторы находятся в ионных каналах, в
течение миллисекунд они повышают проницаемость каналов для Na+, K+ и Са2+ (через один канал
мембраны скелетной мышцы проходит 5 — 107 ионов натрия за 1 с).