

Гликолитическое расщепление глюкозы
Включает в себя 3 стадии: гликолиз‚ окислительное декарбоксилирование (цикл Кребса)‚ электронотранспортная цепь.
Гликолиз
Гликолиз — это процесс генерации энергии в клетке, происходящий без поглощения О2 и выделения СО2. Поэтому его скорость трудно измерить. Основной функцией гликолиза наряду с образованием энергии служит активирование гексоз, достаточно инертных, малореакционноспособных. В результате синтезируются активные молекулы кислоты, подвергающиеся дальнейшим преобразованиям. Кроме того, при гликолизе образуются интермедиаты, необходимые для синтетических процессов в клетке (например, из ФЭП образуются фенольные соединения и лигнин). Важную роль гликолиз играет также при расщеплении крахмала хлоропластов до экспортных продуктов — триоз. Гликолиз осуществляется по схеме‚ представленной на рис. 19.
Суммарное уравнение гликолиза:
С 6Н12О6 2 ПВК + 2 НАД. Н2 + 2 АТФ
Дальнейшая судьба ПВК определяется условиями. При недостатке кислорода ПВК подвергается брожению. При наличии кислорода ПВК направляется в митохондрии‚ где участвует в реакциях цикла Кребса.
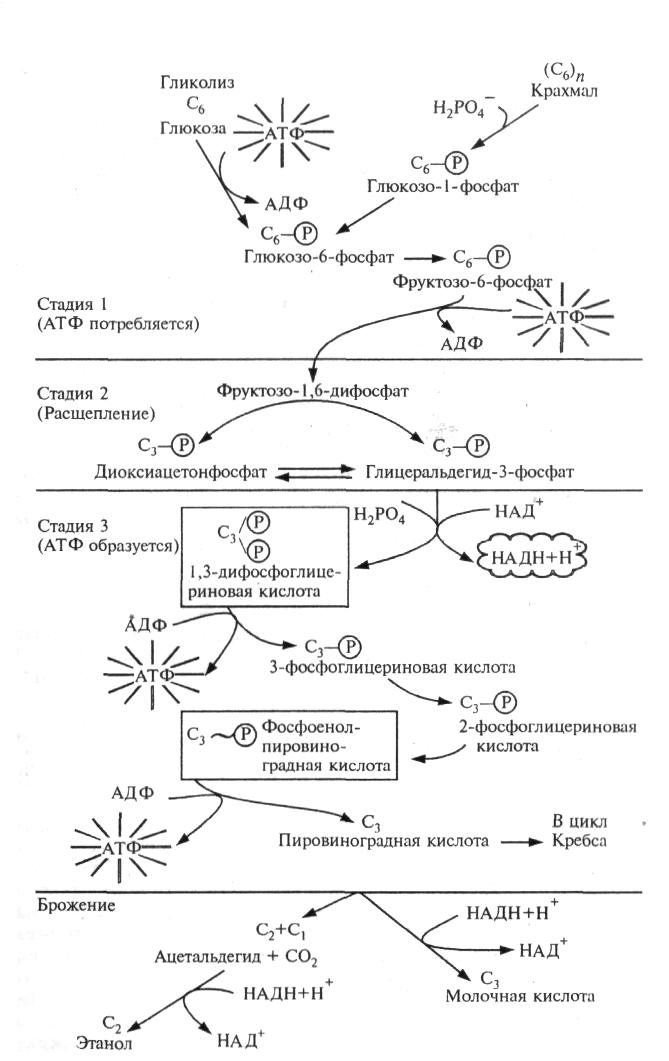
Рис. 19. Схема гликолиза
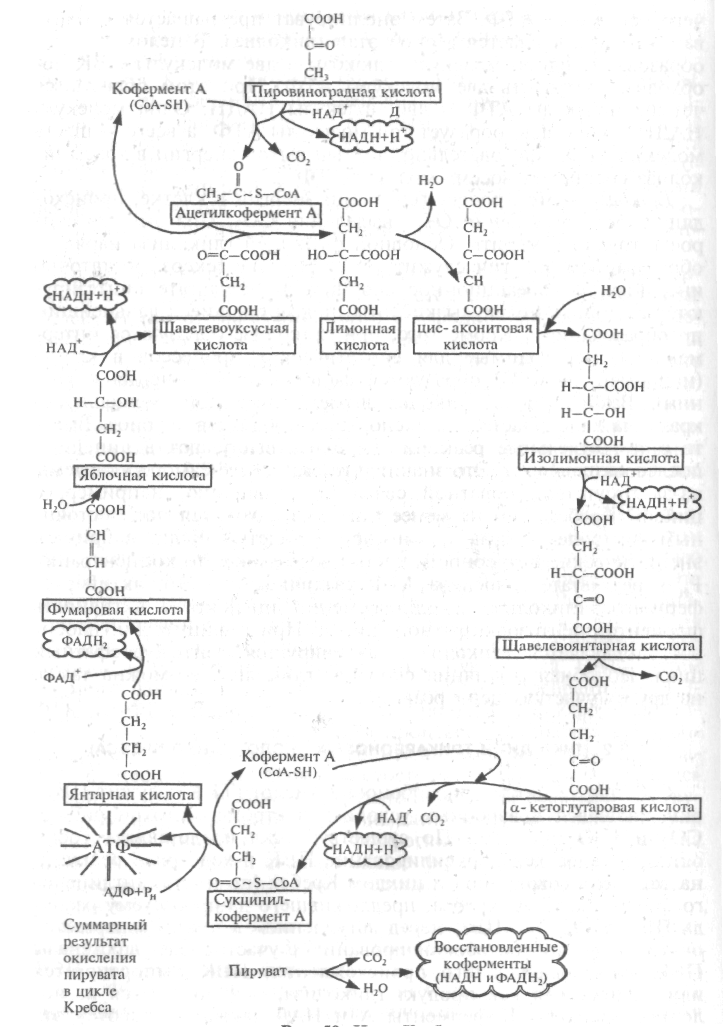
Рис. 20. Цикл Кребса
Цикл Кребса
Субстратом цикла трикарбоновых кислот (ТКК) служит ПВК, окисляющаяся в аэробных условиях в строме митохондрий до СО2 и Н2О (рис. 20). По существу, цикл представляет собой окислительное декарбоксилирование ПВК в ходе ряда реакций, названных в совокупности циклом Кребса (по имени английского биохимика Г.А. Кребса, предложившего данную схему распада ПВК в 1937 г.).
Суммарное уравнение цикла Кребса:
2 ПВК + 6 Н2О 6 СО2 + 8 НАДН2 + 2 ФАДН2
Электрон-транспортная цепь
В рассмотренных реакциях цикла Кребса и при гликолизе молекулярный кислород не участвует. Потребность в кислороде возникает при окислении восстановленных переносчиков НАДН2 и ФАДН2, в которых заключена значительная энергия. Процесс высвобождения энергии, осуществляемый постепенно, заключается в передаче электронов от НАДН2 и ФАДН2 по ЭТЦ на свободный кислород, который, таким образом, служит терминальным акцептором электронов (рис. 61). ЭТЦ растений состоит из четырех мультибелковых комплексов и двух небольших компонентов — убихинона и цитохрома с, расположенных как на внутренней мембране митохондрий, так и внутри нее. Комплексы I, III и IV представляют собой пункты сопряжения транспорта электронов с фосфорилированием. Компоненты дыхательной цепи (переносчики электронов) в определенной мере мобильны, поскольку они как бы представляют собой составную часть жидкого липидного бислоя и, по данным Б. Чанса и др. (1956), расположены в порядке, указанном на рисунке 21.
ЭТЦ представляет сложное образование, состоящее из нескольких десятков белковых молекул и лежащее между метаболитами цикла ТКК с одной стороны и кислородом воздуха с другой. Белки выполняют в основном структурную функцию, входят в состав сопрягающего фактора, а также являются переносчиками электронов. Простетическими группами белков-переносчиков служит наряду с флавином и гемом железо, имеющее переменную валентность и, следовательно, легко переходящее из одной формы в другую. Например, в гемсодержащих цитохромах поступающие к ним по ЭТЦ электроны восстанавливают железо, переводя его из окисной формы (Fe3+) в закисную (Fe2+). Каждый такой переносчик
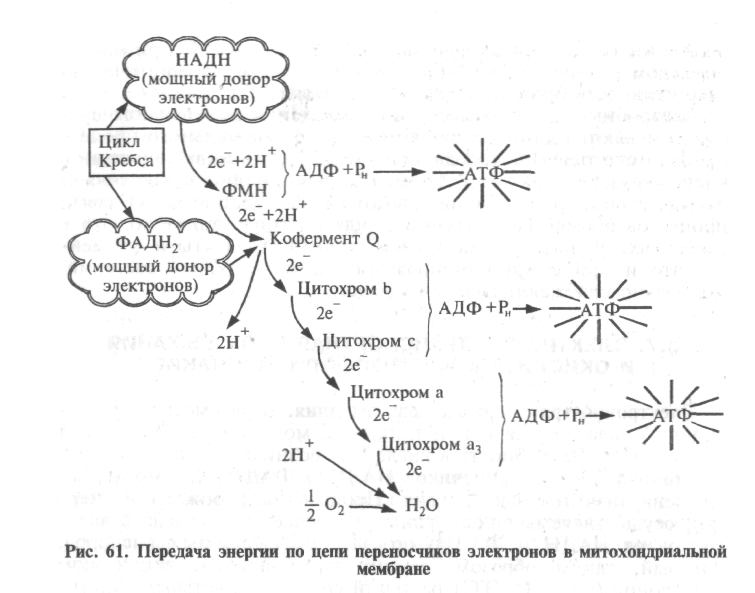
Рис. 21. Электрон-транспортная цепь
находится на несколько более низком уровне восстановленности, чем его предшественник, и в соответствии с этим содержит меньше энергии. Зная окислительно-восстановительный потенциал отдельных компонентов ЭТЦ, их можно расположить в порядке его убывания — от —0,32 у дыхательного субстрата до +0,82 у кислорода.
Электроны скатываются с этой «энергетической» горки благодаря разности потенциалов не сразу, а постепенно, что позволяет, во-первых, избежать неэффективного выброса энергии и, во-вторых, связать энергию в форме химических связей АТФ, образуемых из АДФ и Рн. В этом, по существу, и заключается физиологический смысл транспорта электронов.
На каждую молекулу НАДН2, передающую свои электроны в ЭТЦ, синтезируются три молекулы АТФ, а на каждую молекулу ФАДН2 — две. Процесс фосфорилирования АДФ с образованием АТФ, сопряженный с транспортом электронов от дыхательного субстрата к О2, получил название окислительного фосфорилирования. Этот процесс является основным источником энергии АТФ в клетке в нормальных физиологических условиях, требующим достаточно сложной структуры как ЭТЦ, так и самих митохондрий.