

Цитокинины
Вещества, необходимые для индукции деления растительных клеток, получили название цитокининов. Впервые в чистом виде фактор клеточного деления был выделен из автоклавированного препарата ДНК спермы сельди (К. Миллер, Ф. Скуг и др., 1955). Это вещество было идентифицировано как 6-фурфуриламинопурин (кинетин). Природный цитокинин из незрелых зерновок кукурузы — зеатин — в 1963 г. получил
Д. Летам:
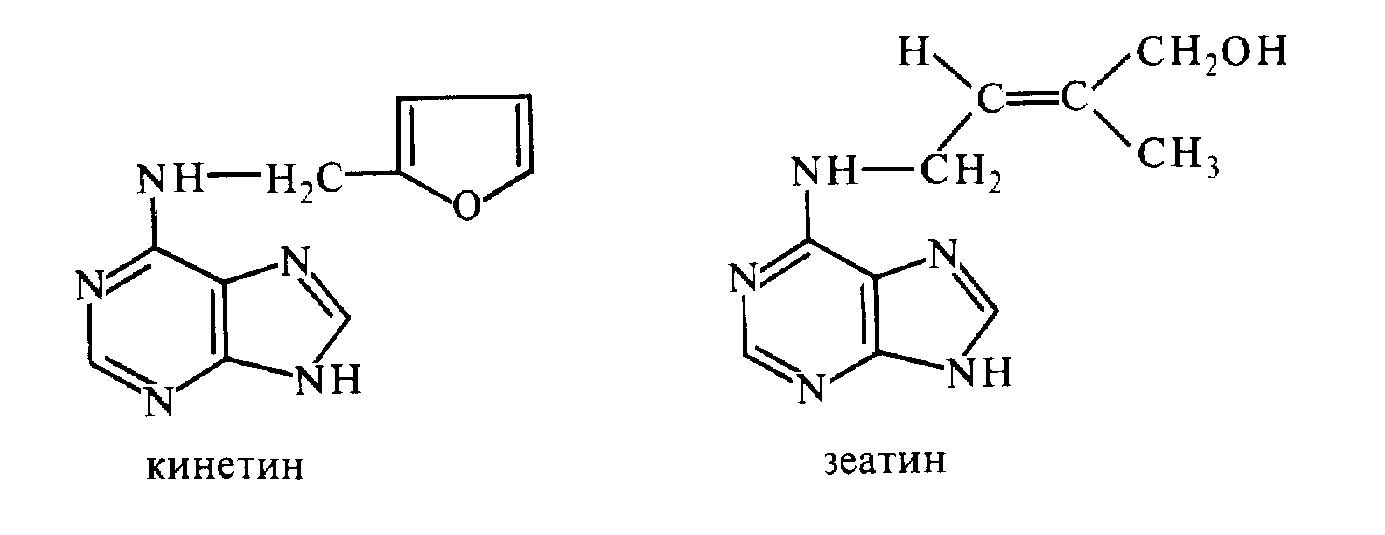
В настоящее время цитокинины обнаружены у микроорганизмов, водорослей, папоротников, мхов и других высших растений. Наиболее богаты ими развивающиеся семена и плоды, а также меристематически активные участки. Установлено, что основное место синтеза цитокинина у вегетирующих растений — апикальные меристемы корней. Из корней цитокинины пассивно транспортируются в надземные органы по ксилеме.
Все природные цитокинины — производные изопентениладенина.
Цитокинины индуцируют деление клеток, однако это их действие наблюдается лишь в присутствии ауксина. Обработка цитокинином вместе с ИУК побуждает дифференцированные клетки растений снова перейти к делению. У семядолей тыквы и листьев фасоли цитокинин активирует рост клеток растяжением. В культуре каллусной ткани цитокинин при повышенных концентрациях (0,5 — 1 мг/л) индуцирует образование побегов, у протонемы мха он способствует развитию гаметофора.
К.Мотес и сотрудники показали, что обогащение тканей цитокинином предотвращает распад хлорофилла и деградацию внутриклеточных структур у изолированных листьев. Эти же авторы установили, что участок изолированного листа, обработанный цитокинином (кинетином), становится зоной притяжения меченых аминокислот и других метаболитов. Таким образом, цитокинин необходим для нормального развития листа и для поддержания его аттрагирующей способности.
На молекулярном уровне цитокинин в комплексе со специфическим белковым рецептором усиливает активность РНК-полимеразы и матричную активность хроматина‚ при этом увеличиваются количество полирибосом и синтез белков, в том числе некоторых ферментов, в частности нитратредуктазы. Имеются данные о действии цитокинина на транспорт К+ Н+ и Са2+.
Гиббереллины
Японский исследователь Е.Куросава в 1926 г. установил, что культуральная жидкость фитопатогенного гриба Gibberella fujikuroi содержит химическое вещество, способствующее сильному вытягиванию стеблей у растений. Т.Ябута (1938) выделил это вещество в кристаллическом виде и назвал его гиббереллином. В 1954 г. англичанин Б.Кросс расшифровал структуру гибберелловой кислоты — тетрациклического дитерпеноида:
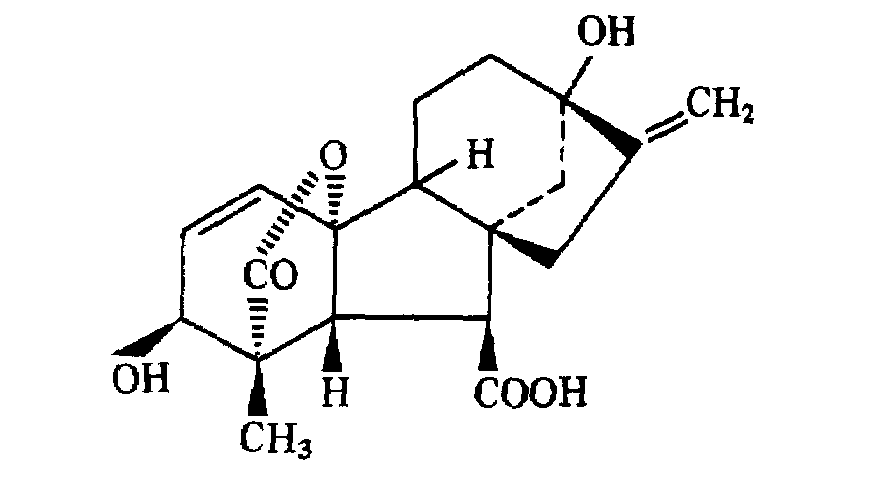
В настоящее время обнаружено более 60 различных гиббереллинов кислой и нейтральной природы. В связи с большим количеством открытых гиббереллинов был введен шифр ГА или просто А. Согласно этой номенклатуре гибберелловая кислота стала обозначаться как ГА3.
ГА и ГА-подобные вещества обнаружены у грибов, водорослей и высших растений. Наибольшее количество гиббереллинов у высших растений содержится в незрелых семенах. Гиббереллины синтезируются главным образом в листьях, а также в корнях. Свет стимулирует образование ГА. Транспорт ГА происходит пассивно с ксилемным и флоэмным током. Как и все полиизопреновые соединения, ГА синтезируются из ацетил-СоА через мевалоновую кислоту и геранилгераниол; ближайший предшественник ГА — каурен. Связанные в виде гликозидов ГА являются запасной и транспортной формами.
Действие ГА на растения наиболее типично проявляется в удлинении их стебля. Особенно наглядно это прослеживается у карликовых растений и у длиннодневных растений, находящихся в фазе розетки. У последних гиббереллин способствует образованию цветоноса, а часто и зацветанию. Места действия ГА — апикальные и интеркалярные меристемы, где под влиянием этого фитогормона активируется деление клеток, ускоряется также растяжение клеток, однако непосредственно на этот процесс ГА действует слабо. Гиббереллины не стимулируют рост корня, а в повышенных концентрациях даже ухудшают его состояние. Как правило, добавки ГА не требуется и при выращивании культуры тканей. Обработка гиббереллином выводит семена и клубни некоторых растений из состояния покоя.
Активирующее действие ГА на синтез нуклеиновых кислот и белков было выявлено у многих растительных объектов. Наиболее важные результаты были получены при использовании алейронового слоя эндосперма зерновок ячменя. В этом объекте пол действием гиббереллина индуцируется синтез матричных РНК, которые кодируют образование α-амилазы и некоторых других гидролаз. Как и другие фитогормоны, ГА прежде всего взаимодействует с цитоплазматическим рецептором белковой природы. Показано также, что гиббереллин активирует ферменты, ответственные за синтез фосфолипидов, входящих в состав мембран, образование гранулярного ЭР и секрецию ферментов. В тканях, обработанных гиббереллином‚ как правило‚ возрастает содержание ИУК.