

67. Расхождение бактериальных хромосом.
МИТОЗ, способ деления клеток, при котором генетический материал (хромосомы) распределяется поровну между новыми (дочерними) клетками. Начинается с разделения ядра на два дочерних. Аналогично делится и цитоплазма. Процессы, происходящие от одного деления до другого, называются митотическим циклом. Он состоит из 2 стадий – интерфазы (стадии покоя) и собственно митоза (стадии деления). В интерфазе в клетке происходит образование ДНК. Интерфаза делится на 3 периода. В первый период, продолжающийся 12–24 ч, происходит накопление РНК и белков. Второй период (синтетический) характеризуется образованием ДНК, в результате чего её количество удваивается. В течение третьего периода (постсинтетического) происходит накопление энергии, после чего клетка из стадии интерфазы переходит к митозу. Митоз проходит 4 последовательные фазы – профазу, метафазу, анафазу и телофазу. В профазе хромосомы уплотняются, скручиваются в спирали и становятся видимыми под микроскопом. Мембрана ядра растворяется под действием ферментов, ядрышко исчезает. Центриоли начинают расходиться к полюсам. Между полюсами формируется веретено деления клетки – структура, состоящая из РНК и белка. К концу профазы хромосомы удваиваются, но члены каждой пары удерживаются рядом. В метафазе они располагаются по экватору клетки. Хроматиды прикреплены к нитям веретена и уже начинают отсоединяться. В анафазе каждая хроматида приобретает собственную центромеру, удлиняется и становится дочерней хромосомой. Нити веретена, прикреплённые к центромерам, разводят «молодые» хромосомы к полюсам клетки. В телофазе дочерние хромосомы достигают полюсов, их спирали раскручиваются, удлиняются и опять становятся плохо видимыми в микроскоп. Образуется ядерная оболочка, вновь появляется ядрышко. В результате клетка имеет двойное количество клеточных структур и общую цитоплазму. В конце митоза происходит её деление. В экваториальной зоне клетки образуется перетяжка, делящая её на 2 дочерние. У растений на месте перетяжки образуется пластинка из целлюлозы.
68. Схема митотического деления клетки
69,70 Регуляция синтеза белка путём индукции и репрессии
Бактериальная клетка способна запустить или прекратить синтез того или иного фермента в зависимости от присутствия соответствующего субстрата. Для этого бактериальные гены объединены в группы ( кластеры ) таким образом, что все ферменты, необходимые для осуществления определенного пути биосинтеза, детерминируются генами, сцепленными друг с другом. Вся группа генов может транскрибироваться в одну полицистронную мРНК, которая последовательно транслируется рибосомами с образованием каждого из белков. Такая форма организации позволяет координировано регулировать выражение всех генов одной единицы транскрипции. Экспрессия генов у прокариот регулируется главным образом на уровне транскрипции. Роль сигнальных веществ для запуска транскрипции играют молекулы-эффекторы , представляющие собой низкомолекулярные соединения, которые являются либо субстратом для фермента, либо продуктом ферментативной деятельности соответственно. Индукция и репрессия представляют собой разные стороны одного и того же явления. Малые молекулы, индуцирующие образование ферментов, способных метаболизировать их, называются индукторами . Те же, которые предотвращают образование ферментов, способных синтезировать их, - корепрессорами . Молекулы-эффекторы не могут вступать в прямое взаимодействие с ДНК, посредником для них служит специальный регуляторный белок . Регуляторный белок, который связывается с ДНК в отсутствии индуктора, называется репрессором .
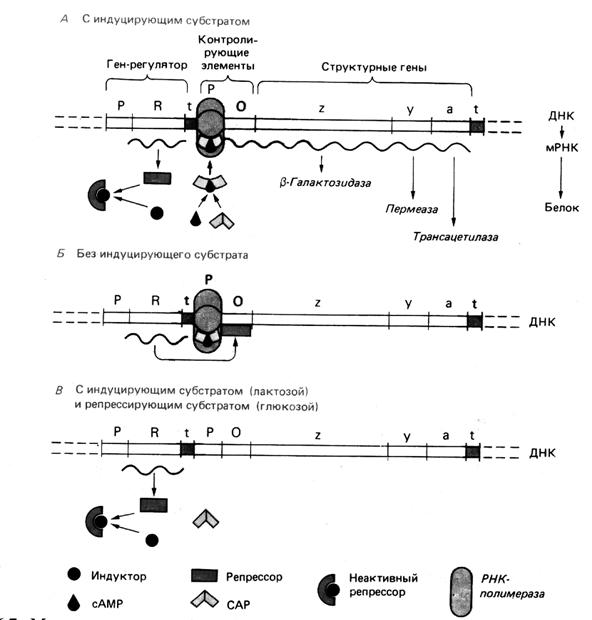
Рис. 2. Модель регуляции лактазного оперона, активность которого определяется как индуцирующим действием субстрата, так и катаболитной репрессией. Для транскрипции оперона необходимо присоединение САР (сАМР-рецепторного белка) к промотору. Оно происходит только в присутствии сАМР. Глюкоза тормозит синтез сАМР и тем самым транскрипцию lac -оперона. (Шлегель Г., 1987).
За синтез регуляторных белков ответственны регуляторные гены . В присутствии белка-репрессора транскрипция блокирована; его удаление обусловливает доступ РНК-полимеразы к генам и запуск транскрипции. Прекращение синтеза фермента при помощи белка-репрессора получило название репрессии . Репрессия позволяет бактериальной клетке избежать перевода своих ресурсов на ненужную в данный момент синтетическую активность. Если индуктор присутствует в клетке в высокой концентрации, то в результате специфического присоединения к регуляторному белку он изменяет его конформацию и тем самым - его способность связываться с ДНК. Контроль транскрипции достигается взаимодействием регуляторного белка с регуляторным сайтом, называемым оператором , который расположен между структурными генами и промотором (участком, распознаваемым ДНК-зависимой РНК-полимеразой). Промотор служит местом связывания РНК-полимеразы, и от него начинается транскрипция. Совокупность промотора, оператора и структурных генов образует оперон . Оперон является функциональной генетической единицей, регулирующей экспрессию одного или группы генов (рис. 2).
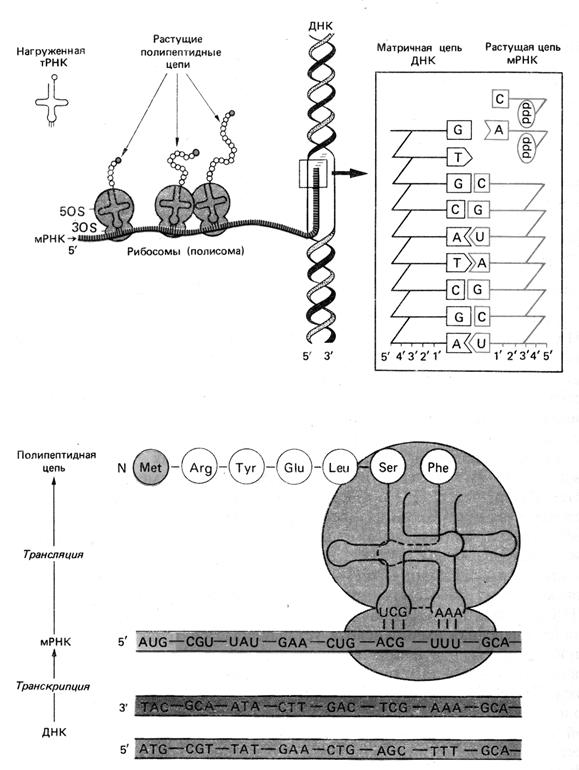
Рис. 1. Биосинтез белка. Перенос генетической информации осуществляется в два этапа. Сначала на матричной цепи ДНК образуется мРНК. Затем во время перемещения рибосомы вдоль мРНК (на схеме - слева направо) различные тРНК подводят к ней аминокислоты и устанавливают их в положение, определяемое триплетами мРНК. Аминокислоты соединяются между собой пептидными связями. (Шлегель Г., 1987).
Так растет
полипептидная цепь по мере продвижения
рибосомы вдоль мРНК. Одновременно
происходит закручивание этой цепи и
свертывание ее в клубок, определяемое
последовательностью аминокислот и
природой их боковых цепей (гидрофобные
и гидрофильные группы), и в результате
возникает структура, обусловливающая
специфические свойства и функцию данного
белка. К мРНК обычно прикрепляется
несколько рибосом, так что на одной и
той же матрице одновременно синтезируется
несколько полипептидных цепей. На конце
мРНК находится кодон, от которого зависит
отделение сформированной полипептидной
цепи от рибосомы (рис. 1).
Таким образом,
нуклеотидная последовательность ДНК
представляет собой закодированную
«инструкцию», определяющую структуру
специфического белка. Этот универсальный
процесс передачи информации при
репликации ДНК, транскрипции и трансляции
применим как к эукариотам ипрокариотам.