

3. Схема дуги рефлекса растяжения (миотатического рефлекса):
1 – α-мотонейрон; 2 – нейрон, расположенный в передних рогах спинного мозга (клетка Реншоу); 3 – сухожильный рецептор; 4 – мышечное веретено со спиралевидным рецептором; 5 – γ-мотонейрон; 6 – чувствительные клетки спинномозгового узла; 7 – экстрапирамидные проводники к γ-мотонейрону; 8 – пирамидная система.
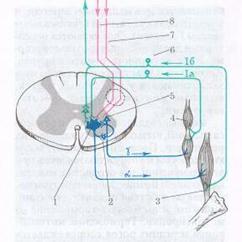
Рефлекс растяжения мышцы обеспечивает сохранение положения тела благодаря поддержанию мышечного тонуса. Он также предотвращает повреждение, давая возможность мышце отвечать на внезапное или неожиданное увеличение длины. Механизм рефлекса следующий:
1. Когда мышца удлиняется, мышечные веретена также растягиваются, заставляя каждое веретено посылать нервный импульс спинному мозгу.
2. При получении импульса спинной мозг немедленно посылает импульс обратно растянутым мышечным волокнам, заставляя их сокращаться, чтобы сопротивляться дальнейшему мышечному растяжению. Этот циркулярный процесс называется рефлекторная дуга.
3. Импульс одновременно посылается от спинного мозга до антагониста мышечного сокращения (т. е. мышце, выступающей против сокращения), вызывая расслабление антагониста, чтобы он не мог сопротивляться сокращению растянутого мышечного волокна. Этот процесс называется реципрокное торможение.
4. Параллельно этому спинальному рефлексу нервные импульсы также посылаются от спинного к головному мозгу с целью передачи информации относительно длины мышцы и скорости ее сокращения. Головной мозг посылает обратно нервные импульсы к мышце для поддержки соответствующего мышечного тонуса, чтобы обеспечить требуемое положение тела и движения.
5. Тем временем чувствительность к растяжению мелких интрафузальных мышечных волокон в пределах мышечного веретена сглаживается и регулируется гамма-эфферентными нервными волокнами, отходящими от двигательных нейронов в спинном мозге. Таким образом, гамма-мотонейрон-рефлекторная дуга обеспечивает равномерность сокращения мышечной ткани, которое иначе было бы хаотичным, если бы мышечный тонус основывался на одном рефлексе растяжения.
4. Выделяют 5 главных ритмов, имеющих различные частотный диапазон, амплитуду и функциональное значение.
1. Дельта-ритм (0,5-4 Гц). Возникает при естественном и наркотическом сне, а так же наблюдается при регистрации ЭЭГ от участков коры, граничащих с областью, пораженных опухолью. У здорового взрослого человека практически отсутствует.
2. Тэта-ритм (5-7 Гц). Наиболее выражен в гиппокампе. Связан с поисковым поведением, усиливается при эмоциональном напряжении. Его иногда так и называют стресс-ритм. Сопровождается переживанием как положительных, так и отрицательных эмоций.
3. Альфа-ритм (8-13 Гц). Основной человеческий ритм, наблюдающийся в состоянии бодрствования, медитации и длительной монотонной деятельности. Наиболее выражен в затылочных областях головного мозга. Обычно эти колебания испытывают определенную модуляцию, так называемое «веретено альфа-ритма» – постепенное увеличение, а затем уменьшение амплитуды колебания альфа-ритма. Длительность веретен доля секунды-несколько секунд. «Десинхронизация» альфа-ритма – реакция пробуждения на раздражитель.
4. Мю-ритм — по частотно-амплитудным характеристикам сходен с альфа-ритмом, но преобладает в передних отделах коры больших полушарий.
5. Бета-ритм (15-35 Гц) – локализуется в предцентральной и фронтальной коре. Наблюдается при умственной активности у взрослых, причем значимое усиление высокочастотной активности наблюдается при умственной деятельности, включающей элементы новизны, в то время как стереотипные, повторяющиеся умственные операции сопровождаются ее снижением. Переход к состоянию напряжения сопровождается появлением бета-активности.
6. Гамма-ритм (выше 35 Гц). В предцентральной, теменной, височной зонах коры. Наблюдается при решении задач, требующих максимального сосредоточения.
ЭЭГ-характеристики спокойного бодрствования определяются как индивидуальными особенностями каждого человека, так и состоянием зрительного анализатора. Вследствие этого следует выделять состояние покоя с закрытыми и открытыми глазами.
Доминирующей в электрограммах спокойного бодрствования с закрытыми глазами более чем 85% людей является альфа-активность . Она может быть как модулирована , так и не модулирована по амплитуде. Состояние покоя с открытыми глазами характеризуется, прежде всего, некоторым снижением амплитуды доминирующих в ЭЭГ альфа-колебаний . Как правило, это снижение наиболее выражено в ЭЭГ затылочных отведений, непосредственно связанных со зрительной функцией. Это приводит к нарушению характерного для состояния покоя с закрытыми глазами градиента амплитуд.
5. При локальных поражениях спинного мозга человека можно наблюдать исчезновение различных рефлексов в зависимости от локализации участка поражения. Так, при поражении нескольких грудных сегментов спинного мозга наблюдается выпадение потоотделения и сосудодвигательных реакций и потеря чувствительности кожных покровов в соответствующих метамерах груди и живота, а также двигательный паралич отдельных мышечных групп. Подобные многочисленные наблюдения свидетельствуют об относительно сегментарном расположении спинномозговых центров. Отмечая сегментарную локализацию ряда спинномозговых центров, надо подчеркнуть, что в спинном мозгу имеются многочисленные межсегментарные связи, обеспечивающие функциональное единство всего спинного мозга.
13
1. Торможение — в физиологии — активный нервный процесс, вызываемый возбуждением и проявляющийся в угнетении или предупреждении другой волны возбуждения. Обеспечивает (вместе с возбуждением) нормальную деятельность всех органов и организма в целом. Имеет охранительное значение (в первую очередь для нервных клеток коры головного мозга), защищая нервную систему от перевозбуждения.
Постсинаптическое — основной вид первичного торможения, вызывается возбуждением клеток Реншоу и вставочных нейронов. При этом типе торможения происходит гиперполяризация постсинаптической мембраны, что и обуславливает торможение. Примеры первичного торможения:
Возвратное — нейрон воздействует на клетку, которая в ответ тормозит этот же нейрон.
Реципрокное — это взаимное торможение, при котором возбуждение одной группы нервных клеток обеспечивает торможение других клеток через вставочный нейрон.
Латеральное — тормозная клетка тормозит расположенные рядом нейроны. Подобные явления развиваются между биполярными и ганглиозными клеткамисетчатки, что создает условия для более четкого видения предмета.
Возвратное облегчение — нейтрализация торможения нейрона при торможении тормозных клеток другими тормозными клетками.
Постсинаптическое торможение(лат. post позади, после чего-либо + греч. sinapsis соприкосновение, соединение) - нервный процесс, обусловленный действием на постсинаптическую мембрану специфических тормозных медиаторов (глицин, гаммааминомаслянная кислота), выделяемых специализированными пресинаптическими нервными окончаниями. Медиатор, выделяемый ими, изменяет свойства постсинаптической мембраны, что вызывает подавление способности клетки генерировать возбуждение. При этом происходит кратковременное повышение проницаемости постсинаптической мембраны к ионам К+ или CI, вызывающее снижение ее входного электрического сопротивления и генерацию тормозного постсинаптического потенциала (ТПСП). Возникновение ТПСП в ответ на афферентное раздражение обязательно связано с включением в тормозной процесс дополнительного звена - тормозного интернейрона, аксональные окончания которого выделяют тормозной медиатор. Специфика тормозных постсинаптических эффектов впервые была изучена на мотонейронах млекопитающих (Д. Экклс, 1951). В дальнейшем первичные ТПСП были зарегистрированы в промежуточных нейронах спинного и продолговатого мозга, в нейронах ретикулярной формации, коры больших полушарий, мозжечка и таламических ядер теплокровных животных.
2. Все спинальные рефлексы можно объединить в две группы по следующим признакам. Во-первых, по рецепторам, раздражение которых вызывает рефлекс: а) проприорецептивные, б) висцероре-цептивные и в) кожные рефлексы. Последние являются защитными. Рефлексы, возникающие с проприорецепторов, участвуют в формировании акта ходьбы и регуляции мышечного тонуса. Висце-рорецептивные рефлексы возникают с интерорецепторов (рецепторов внутренних органов) и проявляются в сокращениях мышц передней брюшной стенки, грудной клетки и разгибателей спины. Во-вторых, спинальные рефлексы целесообразно объединить по органам (эффекторам рефлекса): а) рефлексы конечностей, б) брюшные, в) органов таза. рефлексы конечностей: сгибательные, разгибательные, ритмические и рефлексы позы.
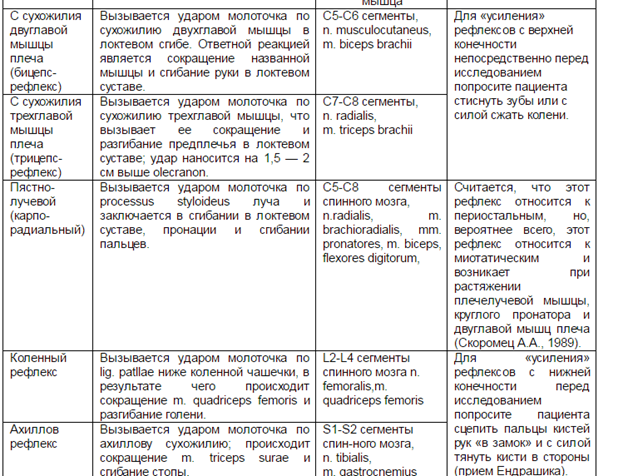
3. Важную роль в управлении движениями выполняет мозжечок. Он обеспечивает сохранение равновесия, поддержание позы, регуляцию и перераспределение мышечного тонуса, тонкую координацию движений.
Функция палеоцеребеллума или средней части - это взаимная координация позы и целенаправленного движения, а также координация выполнения сравнительно медленных движений на основе механизма обратной связи. Эта функция реализуется с участием двух ядер мозжечка - пробковидного и шаровидного. Они влияют на деятельность красного ядра и РФ продолговатого мозга. Эта часть мозжечка работает на основании информации, поступающей от мышечных рецепторов и двигательной коры. Функция коррекции медленных движений имеет большое значение в процессе обучения, но она не может использоваться при выполнении быстрых и очень сложных движений.
Неоцеребеллум или латеральная часть мозжечка - играет важную роль в программировании сложных движений, выполнение которых идёт без использования механизма обратных связей. Информация в неоцеребеллум поступает от ассоциативных зон коры (формирующих замысел движения). Вначале она доставляется в нейроны моста и оттуда уже поступает в неоцеребеллум. От нейронов коры мозжечка информация идет на зубчатое ядро, а от него направляется через таламус к двигательной коре, откуда она поступает к красному и вестибулярному ядру, и по пирамидному пути к альфа-мотонейронам спинного мозга. В итоге становится возможным выполнение целенаправленного движения, выполняемого с большой скоростью, при этом сохраняется равновесие тела.
4. Таламус, или зрительный бугор, является одним из важнейших образований головного мозга, он тесно связан морфологически и функционально с корой больших полушарий. Не без основания считается, что в таламусе хранятся ключи к тайнам коры головного мозга. Деятельность таламуса тесно связана с анализом афферентных сигналов, организацией интегратив-ных процессов, он участвует в механизмах регуляции функционального состояния и высшей нервной деятельности.
Специфические, или релейные переключательные, ядра таламуса характеризуются следующими особенностями.
Имеют локальную проекцию в строго определенные участки коры.
Моносинаптически связаны аксосоматическими синапсами с нейронами третьего-четвертого слоев коры.
В них переключается афферентная импульсация от периферических рецепторов или от первичных воспринимающих ядер нижележащих стволовых структур, а также от внесенсорных источников. Основную массу клеток составляют «релейные»
нейроны с небольшим диаметром тела, длинным аксоном и слаборазвитой дендритной системой. «Релейные* клетки окружены вставочными интернейронами, многие из которых имеют тормозную функцию. Содержатся также длинноаксонные клетки, дающиеколлатерали в ретикулярную формацию среднего мозга, и неспецифические ядра таламуса, которые выполняют ассоциативную или интегративную функции. Одиночное раздражение соответствующей рецепторной области вызывает в специфическом ядре первичный положительно-отрицательный электрический ответ, опережающий ответ в коре.
Одиночное раздражение специфического ядра вызывает в соответствующей проекционной зоне корыпервичный положительно-отрицательный ответ, а ритмическая низкочастотная стимуляция — реакцию усиления (Демпси и Моррисон).
К специфическим ядрам таламуса относят переднее дорсальное, переднее вентральное, переднее медиальное, вентральное постлатеральное, вентральное постмедиальное, вентральное латеральное, а также латеральное и медиальное коленчатые тела. В функциональном отношении специфические ядра представляют собой важнейшую часть основных сенсорных и моторных систем, разрушение релейных ядер приводит к полной и необратимой потере соответствующей чувствительности или нарушениям движений.
5. Сокращения скелетных мышц, а также их тонус связаны с возбуждением а-мотонейронов, находящихся в спинном мозге. Сила сокращения мышцы и ее тонус зависят от количества возбужденных мотонейронов и частоты их разрядов.
Нарушение двигательных функций возникают как при повреждении указанных отделов центральной нервной системы, так и при нарушении проведения импульсов по двигательным нервам и передачи импульсов с нерва на мышцу.
Наиболее распространенной формой двигательных нарушений являются паралич и парез - потеря или ослабление движений вследствие нарушения двигательной функции нервной системы. Паралич мышц одной половины тела называется гемиплегией, обоих верхних или нижних конечностей - параплегией, всех конечностей - тетраплегией. В зависимости от патогенеза паралича тонус пораженных мышц может быть либо утрачен (вялый паралич), либо повышен (спастический паралич). Кроме того, различают паралич периферический (если он связан с повреждением периферического мотонейрона) и центральный (в результате поражения центральных двигательных нейронов).
14
1) Механизм возбуждения в синапсах цнс Под влиянием нервных импульсов, приходящих к окончанию аксона, из синаптических пузырьков выделяется медиатор порционно в виде квантов, в каждом из которых находится несколько тысяч молекул. Каждый синаптический пузырек содержит квант медиатора. Для высвобождения медиатора необходимы ионы кальция. Когда нервный импульс доходит до пресинаптической мембраны в результате биохимических реакций активируется ее кальциевые каналы и освобождаются ионы кальция. В присутствии ионов кальция пузырьки с медиатором проходят через пресинаптическую мембрану и выделяются в синаптическую щель. Этот процесс получил название экзоцитоза. После выхода медиатора из пузырька, окружающая его мембрана включается обратно в мембрану пресинаптическую (эндоцитоз). 2) нарисовать схему рефлекторной дуги спинномозгового соматического и вегетативного рефлекса. Указать их различия Морфологические различия: 1) очаговость расположения вегетативных ядер в спинном и головном мозге; 2) широкое ее распространение в организме; 3) отсутствие строгого сегментарного строе ния; 4) наличие многочисленных узлов (ганглиев 1,2,3-его порядков) в составе перифе рической части; 5) вегетатив ные волокна тоньше, чем соматические, и нервные импуль сы по ним передаются с меньшей скоростью. 3) что такое статокинетические рефлексы? Их значение, примеры Статокинетические рефлексы возникают при вращении и при любых перемещениях тела в пространстве независимо от того, активно или пассивно совершаются эти перемещения. Ярким примером подобных рефлексов является лифтный рефлекс: подъем лифта вызывает сгибание ног, остановка — их выпрямление. Морская и воздушная бо-лезни, проявляющиеся в плохом самочувствии, тошноте и т. д., тоже являются статокинетическими рефлексами, но здесь воздействие оказывается главным образом на внутренние органы. Роль тонических рефлексов в двигательной деятельности артистов балета. Тонические рефлексы осуществляются автоматически. Человек не задумывается ни о том, куда надо наклонить голову, ни о том, какие мышцы следует напрячь, для того чтобы сохранить равновесие. Но кора полушарий головного мозга контролирует деятельность отделов мозга, в которых замыкаются дуги тонических рефлексов, управляет ими. Например, усилием воли тонические рефлексы можно затормозить: при разучивании новых движений и комбинаций приходится подавлять тонические рефлексы. Защищая организм от травм, они мешают выполнять новые, непривычные, сложные для него движения. 4) основные ритмы ЭЭГ их характеристика Альфа-ритм — регулярный ритм синусоидальной формы, с частотой 8-13 гц (колебаний в 1 с) и амплитудой 20-80 мкВ (микровольт). Альфа-ритм регистрируется при отведении биопотенциалов от всех зон коры большого мозга, но более постоянно — от затылочной и теменной областей. Альфа-ритм регистрируется у человека в условиях физического и умственного покоя, обязательно при закрытых глазах и отсутствии внешних раздражений. Бета-ритм имеет частоту колебаний 14-35 гц. Этот ритм низкоамплитудный: всего 10-30 мкВ. Он может быть зарегистрирован при отведении потенциалов от любых областей коры большого мозга, но более выражен в лобных долях. При нанесении различных раздражений, открывании глаз, умственной работе альфа-ритм быстро сменяется бета-ритмом. Это явление смены редкого ритма на более частый получило название реакции активации (или десинхронизации). Тета-ритм имеет частоту 4-7 гц, его амплитуда 100-150 мкВ. Он наблюдается в состоянии неглубокого сна, при кислородном голодании организма, при умеренном по глубине наркозе. Дельта-ритм характеризуется медленными колебаниями потенциалов с частотой 0,5-3 гц, амплитуда его высокая: 250-300 мкВ, может доходить до 1000 мкВ. Он обнаруживается при отведении биопотенциалов от всех зон коры большого мозга, во время глубокого сна, а также при наркозе. У детей до 7 лет дельта-ритм может быть зарегистрирован и в бодрствующем состоянии.
15
1) в чем заключается явление суммации в нервных центрах? Виды суммации, примеры Имеется 2 вида суммации: временная и пространственная. Временная — возникает ответная реакция при действии нескольких следующих друг за другом раздражителей. Механизм: суммируются возбуждающие постсинаптические потенциалы рецептивного поля одного рефлекса. Происходит суммация во времени потенциалов одних и тех же групп синапсов. Пространственная суммация — возникновение ответной реакции при одновременном действии нескольких подпороговых раздражителей. Механизм: суммация возбуждающего постсинаптического потенциала от разных рецептивных полей. Суммируются потенциалы разных групп синапсов. 2) статические рефлексы. Сущность, значение, рефлексогенные зоны Статические рефлексы регулируют тонус скелетных мышц с целью удержания определенного положения тела. Статические рефлексы подразделяются на позные, благодаря которым сохраняется вертикальная поза, и установочные рефлексы, про являющиеся при смене одной позы на другую, например при вставании из положения сидя или лежа 3) функциональное значение таламуса Функциональная значимость ядер таламуса определяется не только их проекциями на другие структуры мозга, но и тем, какие струк туры посылают к нему свою информацию. В таламус приходят сигналы от зрительной, слуховой, вкусовой, кожной, мышечной систем, от ядер черепно-мозговых нервов ствола, мозжечка, блед ного шара, продолговатого и спинного мозга. 4) характеристика Альфа и Бетта ритма ЭЭГ и их функциональное значение Альфа-ритм — регулярный ритм синусоидальной формы, с частотой 8-13 гц (колебаний в 1 с) и амплитудой 20-80 мкВ (микровольт). Альфа-ритм регистрируется при отведении биопотенциалов от всех зон коры большого мозга, но более постоянно — от затылочной и теменной областей. Альфа-ритм регистрируется у человека в условиях физического и умственного покоя, обязательно при закрытых глазах и отсутствии внешних раздражений. Бета-ритм имеет частоту колебаний 14-35 гц. Этот ритм низкоамплитудный: всего 10-30 мкВ. Он может быть зарегистрирован при отведении потенциалов от любых областей коры большого мозга, но более выражен в лобных долях. При нанесении различных раздражений, открывании глаз, умственной работе альфа-ритм быстро сменяется бета-ритмом. Это явление смены редкого ритма на более частый получило название реакции активации (или десинхронизации).