

Синтез белка
Реализация содержащейся в ДНК наследственной информации осуществляется в процессе синтеза белка. ДНК преимущественно находится в ядрах, синтез белка осуществляется в рибосомах, расположенных в цитоплазме. Передача информации от ДНК к рибосоме осуществляется с помощью информационной, или матричной, РНК (и-РНК), которая синтезируется на ДНК по принципу комплементарности с помощью специального фермента из класса синтетаз. Триплет РНК, соответствующий' одной аминокислоте, называется кодоном.
Из ядра и-РНК поступает в цитоплазму и здесь последовательно вступает в контакт с несколькими рибосомами. Группа рибосом, одновременно расположенных на одной и-РНК, называется полирибосомой, или полисомой. Рибосомы неспецифичны и потому могут соединяться с любой и-РНК и синтезировать любой белок. В рибосоме, связанной с и-РНК, начинается синтез белка. Этот синтез невозможен без участия еще одной формы РНК—транспортной (т-РНК), называемой еще растворимой. Этим названием подчеркивается недолговечность существования данного вида РНК. Схема синтеза белка показана на рис. 18. и-РНК имеет на спирали участок, состоящий из триплета нуклеотидов, комплементарны кодону в и-РНК. Это антикодон. Каждой аминокислоте соответствует специфическая т-РНК, но для того чтобы они соединились, аминокислота должна быть активирована энергией от АТФ.
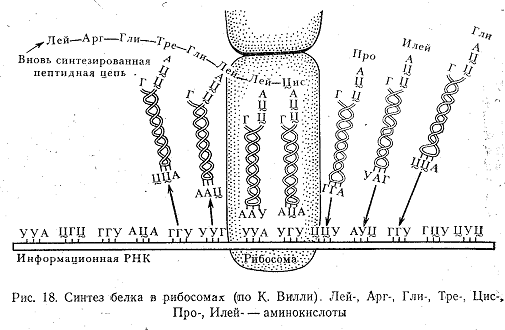
Транспортная РНК, несущая аминокислоту, входит в рибосому. Функция рибосомы, по-видимому, заключается в том, чтобы обеспечить правильное взаимное расположение и-РНК, т-РНК и аминокислоты. Если кодон соответствует антикодону, то т-РНК оставляет аминокислоту на образующейся полипептидной цепочке и покидает рибосому. Рибосома при этом продвигается по нити и-РНК на один триплет. Если же антикодон кодону не соответствует, то т-РНК покидает рибосому вместе с аминокислотой, а рибосома задерживается на непрореагировавшем кодоне и-РНК до тех пор, пока не будет присоединена к синтезирующемуся полипептиду соответствующая этому кодону аминокислота. Так рибосома продвигается по и-РНК от начала до конца нити, и когда она «соскакивает» с и-РНК, молекула белка оказывается завершенной. Одновременно на одной молекуле и-РНК помещается несколько рибосом и синтезируется несколько молекул белка. Период полураспада одной молекулы РНК варьирует от нескольких минут у некоторых микроорганизмов до 12—16 ч у млекопитающих. Хотя и-РНК служит матрицей для синтеза многих молекул, в конце концов она разрушается и должна быть заменена новой. Если бы и-РНК не распадалась, в клетке непрерывно шел бы синтез одних и тех же белков. Распад и-РНК дает клетке возможность производить замену и-РНК и, следовательно, переключаться на синтез нужных в данный момент белков.
Мутагенез
Механизм редупликации ДНК, строгая последовательность присоединения аминокислот при синтезе белковой молекулы, системы регуляции клетки, обеспечивающие синтез определенных белков,— все это, казалось бы, должно гарантировать постоянство наследственных признаков при передаче их потомству. Если бы действительно это было так, то все отличие потомков от предков должно было бы заключаться в перераспределении родительских признаков между детьми или в проявлении влияния внешней среды. Однако накопленный многовековой опыт показывает, что это не так. У потомков появляются признаки, отсутствовавшие у их родителей. Такие изменения, если они способны передаваться потомству, называются мутациями.
Одни из вновь появившихся признаков оказываются полезными организму, другие—вредными. Особи, обладающие полезными при данных условиях признаками, имеют больше шансов оставить потомство. Поэтому в следующем поколении процент особей с такими признаками возрастает. В результате признаки, способствующие процветанию вида, закрепляются в потомстве. Особи с неблагоприятными признаками нередко вообще, не дают потомства.
Для того чтобы признак был наследуемым, он должен быть отражен в строении ДНК.
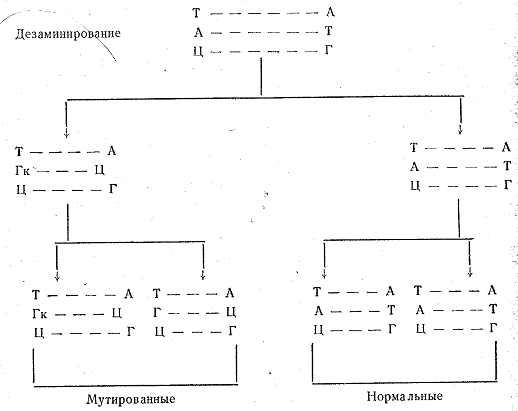
Нарушение строения ДНК при мутагенезе было подтверждено экспериментальными исследованиями. Более того, изучение природы мутационного процесса позволило в некоторых случаях выявить зависимость между характером воздействия и теми изменениями, которые оно вызывает в структуре ДНК. Так, азотистая кислота вызывает дезаминирование нуклеотидных оснований—аденина, цитозина и гуанина. В результате этого аденин (А), например, превращается в гипоксантин (Гк), который в отличие от аденина соединяется не с тимином, а с цитозином. При последующей редупликации цитозин, как обычно, свяжется с гуанином, и в дальнейшем эта замена сохранится. Изменение в составе триплета ДНК приведет к тому, что изменится и кодон в РНК, а это, в свою очередь, повлечет за собой замену одной из аминокислот в молекуле белка.
Например, триплет ТАЦ в ДНК комплементарен кодону АЦГ, который соответствует аминокислоте метионину. Если аденин будет заменен гуанином, то в ДНК образуется триплет ТГЦ, а в РНК—кодон АЦГ, соответствующий уже не метионину, а треонину. Вследствие того что ДНК состоит из двух спиралей нуклеотидов, при замене нуклеотида только в одной спирали мутированную ДНК получит только половина потомков:
Такого рода мутации носят название точковых, или генных. Они могут оказаться полностью или почти безвредными, если при замене нуклеотидов образуется триплет, кодирующий ту же самую аминокислоту или если замена аминокислоты произойдет на менее важном участке молекулы белка, но эти мутации могут сыграть решающую роль, сделав белок-фермент неактивным. Другой вид мутаций заключается в выпадении из спирали ДНК одного нуклеотида. Это приводит к тому, что меняется состав всех последующих триплетов и, следовательно, аминокислот.
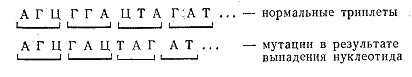
Аналогичный результат получается при удвоении участка какого-либо нуклеотида. Подобные точковые мутации вызываются акридинами, реагирующими с фосфатными группами ДНК наподобие нуклеотидных оснований. Причиной точковых мутаций могут быть различные химические соединения, в том числе азотистые аналоги иприта, эпоксиды, алкилирующие агенты и др. Кроме того, причиной мутации могут быть разнообразные виды радиации: рентгеновские, ультрафиолетовые и гамма-лучи, различные радиоактивные излучения. Вещества, вызывающие мутации, называются мутагенами.
Мутации возникают и спонтанно, без участия мутагенов. Вполне вероятно, что действительное число генных мутаций гораздо больше обнаруживаемых. У человека частота спонтанных мутаций различных генов лежит в пределах от 1·10-5 до 1·10-3 на ген. Всего человек имеет порядка 2,3·104 генов. Следовательно, в среднем в каждом человеке есть один мутировавший ген.
Помимо генных мутаций, существуют хромосомные. Они захватывают более крупные участки ДНК и сопровождаются видимыми изменениями в структуре хромосом или ДНК. Хромосомные перестройки часто бывают следствием воздействия на организмы больших доз излучений.
Многие вещества, содержащиеся в сточных водах различных производств, потенциально мутагенны. Чем больше концентрация мутагенов, тем, естественно, большая вероятность мутаций, но в принципе смертельная мутация может быть вызвана и одной молекулой мутагена, изменившей порядок нуклеотидов в ДНК. По этой причине оценка качества очищенной воды химическими методами может не совпадать с оценкой, сделанной на основании биологического анализа. Это объясняется прежде всего тем, что не все химические соединения, содержащиеся в сточной воде и образовавшиеся в ней, известны. Часто неизвестна их принадлежность к мутагенам. Кроме того, концентрации порядка 0,001 м/г и ниже трудно поддаются анализу, а мутации могут возникнуть и при очень низких концентрациях мутагена.
Мутационный эффект не всегда проявляется в первом поколении, так как он может маскироваться действием здоровых генов. Но и скрытые мутации, постепенно суммируясь в ряде поколений, могут привести к вырождению организмов и отразиться на жизни водоема.
Из всего сказанного следует, что невозможно оценивать качество очищенной воды только по результатам химических анализов. Каждый новый вид очищенных сточных вод и особенно сточных вод химической и лесохимической промышленности должен непременно проходить биологическую проверку.