

Экосистемы анаэробных очистных сооружений
В процессе механической очистки сточных вод значительная часть содержащихся в них загрязнений выпадает в осадок в первичных отстойниках. В процессе биологической очистки образуется активный ил или биопленка, которые выделяются в осадок во вторичных отстойниках. И осадок из первичных отстойников, и активный ил, и биопленка содержат большое количество органических веществ. Дальнейшая переработка осадков осуществляется либо путем сбраживания, либо путем механического обезвоживания без участия микроорганизмов.
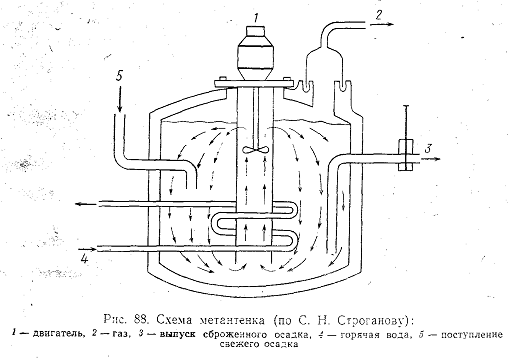
Для сбраживания осадков в настоящее время применяются два основных сооружения: метантенк и камера перегнивания осветлителя-перегнивателя. Биологические процессы в осветлителе-перегнивателе совершенно не изучены, и поэтому в дальнейшем рассматриваются только процессы, совершающиеся в метантенке (рис. 88).
Биологическое изучение анаэробных процессов представляет очень большие трудности (см. гл. IX), и потому чисто инженерные вопросы конструкции и эксплуатации метантенков разработаны намного полнее, чем вопросы исследования сущности биохимических реакций, протекающих в сооружении.
В.процессе превращения веществ в метантенке участвуют как органотрофные, так и литотрофные микроорганизмы. Органотрофы расщепляют углеводы, жиры, белки и другие органические соединения, образуя в качестве конечных продуктов в основном низкомолекулярные жирные кислоты. Эта фаза в работе метантенка называется стадией кислотообразования, или кислым брожением.
Литотрофные микроорганизмы превращают конечные продукты метаболизма органотрофных бактерий в метан и двуокись углерода. Эта фаза называется стадией газообразования, или щелочным брожением.
Обе группы бактерий присутствуют в метантенке одновременно, поэтому кислото- и газообразование осуществляются параллельно. При нормальной работе метантенка появляющиеся при кислом брожении продукты успевают переработаться бактериями второй фазы, вследствие чего в метантенке процесс идет обычно в стадии щелочного брожения.
СТАДИЯ КИСЛОТООБРАЗОВАНИЯ
В стадии кислого брожения в метантенке обнаруживаются разнообразные бактерии, грибы и даже простейшие, попадающие в метантенк с активным илом. Достоверно установлено присутствие в метантенке около 50 видов бактерий, относящихся к 19 родам. Среди них 12 видов бацилл (в том числе такие распространенные формы, как Вас. cereus, Вас. megaterium, Вас. subtilis), 9 видов рода Pseudomonas (в том числе Ps. aeruginbsa), 5 видов Micrococcus, два вида Sarcina vulgaris, Escherichia coil и другие бактерии. Наряду с облигатными анаэробами в стадии кислого брожения присутствуют и факультативно анаэробные и даже аэробные формы, вместе с органотрофами—литотрофы. Бактерии I фазы относятся к различным физиологическим группам. Чаще других встречаются протеолитические аммонифицирующие бактерии, участвующие в разложении белка и аминокислот. Их число достигает 1—3 млн/мл. До 1 млн/мл доходит количество сульфатвосстанавливающих бактерий. Несколько ниже содержание денитрифицирующих и целлюлозоразлагающих бактерий—до 100 тыс/мл. Примерно столько же содержится в осадке бактерий, разлагающих белок с 'образованием сероводорода.
Общее количество аэробных бактерий в осадке, по данным различных авторов, колеблется в пределах от 1 до 130 млн/мл, а анаэробных достигает 15 млрд/мл.
Грибы в стадии кислотообразования представлены очень широко, в метантенке отмечалось 35 родов. Среди них Penicillium, Fusarium, Mucor и др. Отмечено, что внесенные в метантенк дрожжи также приживались в нем. Простейшие в метантенке представлены жгутиковыми, корненожками и ресничными инфузориями, но число их очень невелико. Экологические связи в метантенке и роль отдельных групп микроорганизмов изучены недостаточно.
Основная роль в процессах разложения органических веществ в метантенке принадлежит анаэробным бактериям. Аэробные бактерии, такие, как нитрифицирующие, по-видимому, попадают в метантенк с активным илом и не играют существенной роли в происходящих здесь процессах. То же можно сказать о грибах и простейших.
Понимание сущности процессов в любой экологической системе невозможно без изучения ферментативной активности.
Анализ субстрата в метантенке показал, что в нем содержатся разнообразные ферменты. В осадке обнаружены различные дегидрогеназы, в том числе НАД и НАДФ, кофермент А, киназы, энолазы, лиазы, трансаминазы, ферменты, участвующие в цикле трикарбоновых кислот, а также ферменты из группы цитохромов. Следовательно, химизм анаэробного распада органических соединений во многом подобен аэробному окислению. Основное отличие между ними заключается в том, что конечным акцептором электрона служат органические соединения и двуокись углерода, а не молекулярный кислород.
Кроме окислительно-восстановительных ферментов, в осадке метантенка обнаружены внеклеточные гликолитические ферменты, такие, как протеаза, целлюлаза, амилаза и липаза, расщепляющие соответственно белки, клетчатку, крахмал и жиры.
В результате гидролиза белков с помощью протеаз образуются полипептиды и аминокислоты. Аминокислоты могут разрушаться различными путями в зависимости от того, какие микроорганизмы участвуют в этом процессе, но в конечном счете от аминокислот с помощью ферментов трансаминаз отщепляется аминогруппа, и они превращаются в жирные кислоты.
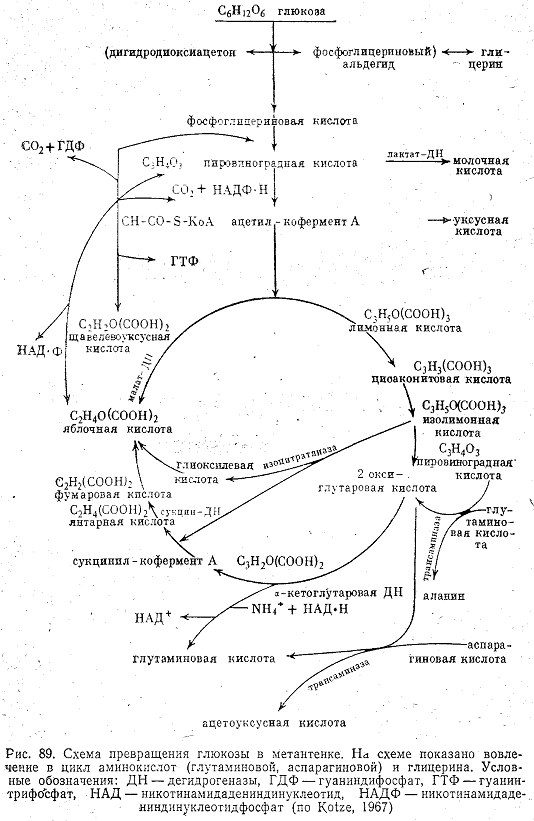
Разрушение жиров в анаэробных условиях происходит путем первоначального расщепления их при помощи липазы на глицерин и жирные кислоты. Затем глицерин, по-видимому, через стадию фосфоглицеринового альдегида превращается в пировиноградную кислоту, а длинные цепочки жирных кислот разрушаются β-окислением6. Химизм этого процесса был экспериментально установлен Мак Карти с сотрудниками (1962) с применением изотопа 14С на примере октановой и пальмитиновой кислот. Конечным акцептором электрона при анаэробном окислении жирных кислот является двуокись углерода. Жирорасщепляющие бактерии не способны образовывать СH4, и при восстановлении СО2 образуется ацетат. Предполагают, что при этом может выделяться газообразный водород, но экспериментального подтверждения пока не получено.
Превращение углеводов в анаэробных условиях также изучалось с применением изотопа 14С. Схема расщепления глюкозы, предложенная Кютц (Kotze) с сотрудниками (1967), представлена на рис. 89. В результате разрушения углеводов тоже образуются кислоты жирного ряда.
Таким образом, конечные продукты стадии кислотообразования—жирные кислоты, двуокись углерода и аммоний с сероводородом, отделившиеся от аминокислот. Наиболее частыми конечными продуктами распада являются уксусная и муравьиная кислоты, в больших количествах обнаруживаются пропионовая и масляная кислоты. Не доказано, но предполагается образование молочной и янтарной кислот. Кроме того, среди конечных продуктов этой стадии обнаружены спирты, кетоны, ацетон, уксусный альдегид.
СТАДИЯ ГАЗООБРАЗОВАНИЯ
Образовавшиеся на первой стадии продукты неполного окисления, такие, как низшие жирные кислоты и низшие спирты, подвергаются дальнейшим превращениям, в результате которых в значительном количестве образуются метан и двуокись углерода. Вследствие разрушения органических кислот реакция среды становится щелочной, и поэтому вторую стадию в метантенке называют щелочным брожением, или стадией газообразования.
Основную роль на стадии газообразования играют метановые или метанообразующие бактерии. Метановые бактерии относятся к строгим анаэробам, изучение их связано с большими трудностями, и потому многие вопросы их физиологии еще недостаточно изучены.
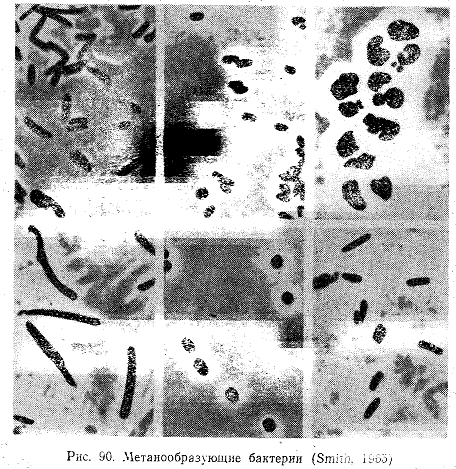
Морфологически метанообразующие бактерии весьма разнообразны. Среди них есть кокки, сарцины, палочки и в соответствии с формой клеток они подразделяются на роды Methanococcus, Methanosarcina, Methanobacterium (рис. 90). Все метановые бактерии не образуют спор.
Физиологически метанообразующие бактерии весьма сходны друг с другом. Они неспособны расти при концентрации кислорода выше 0,1 %. В лабораторных условиях их удается культивировать при окислительно-восстановительном потенциале среды порядка—0,36 В. В естественных условиях метановые бактерии обычно развиваются совместно с другими анаэробными микроорганизмами, и низкий потенциал, необходимый метановым бактериям, создается за счет жизнедеятельности сопутствующей микрофлоры. Метанообразующие бактерии не способны разлагать полимерные соединения и даже углеводы. Для многих из них специфическим субстратом служат продукты брожения.
Процесс образования метана необходим метановым бактериям для получения энергии. Другие способы получения энергии у них неизвестны. Все метановые бактерии могут образовывать метан путем восстановления углекислоты водородом, но многие, кроме того, способны продуцировать метан из простых органических соединений. В зависимости от способности к использованию различных субстратов в качестве источника для образования метана Г. А. Заварзин (1972) предложил подразделить метановые бактерии на четыре группы:
1. Бактерии, использующие только водород и углекислоту или муравьиную кислоту, но не способные образовать метан из органических кислот и спиртов. Они осуществляют только реакцию:
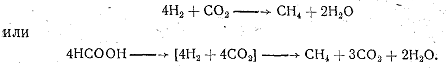
2. Кроме первой реакции, бактерии способны образовывать метан из уксусной кислоты:
![]()
3. Бактерии, кроме первых двух субстратов, способны использовать метанол. Происходит взаимное окисление—восстановление двух молекул спирта:
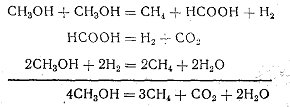
4. Бактерии способны использовать различные органические кислоты. Например:
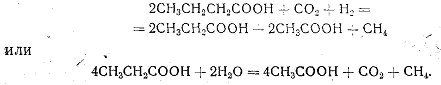
Таким образом, среди метанообразующих бактерий имеются как литотрофные, так и органотрофные организмы.
Метанообразующие бактерии—конкуренты сульфатредуцирующих. Обычно процессы образования сероводорода и метана взаимно исключают друг друга. В метантенках в первую стадию образуется сероводород, а во вторую—метан. Конечным продуктом второй стадии, как видно из приведенных уравнений реакций, наряду с метаном является двуокись углерода.
Суммарный ход
процесса разложения органических
веществ в метантенке показан на рис.
91. Химизму образования метана в последние
годы уделяется большое внимание.
Установлено, что в процессе образования
метана принимают участие фермент
ферредоксин и кофермент А, АТФ и витамин
Bi2. Присутствие таких распространенных
переносчиков водорода, как НАД и НАДФ,
ингибирует метанообразование.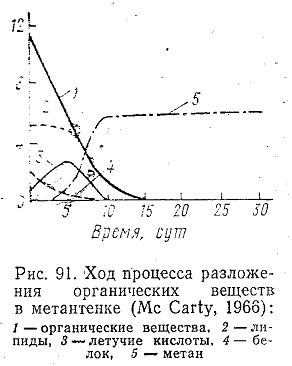
С применением изотопов удалось установить, что 70—75 % метана в метантенках образуется при разложении уксусной кислоты, а 25—30 %—путем синтеза углекислоты и водорода. Количественно водородные метанообразующие бактерии преобладают над бактериями, разлагающими жирные кислоты с образованием
метана. В 1 мл осадка содержится водородных метанообразующих бактерий 107, разлагающих ацетат 105—106, пропионат и бутират 106—107.
Процессы в метантенке, как и все, анаэробные процессы, идут очень медленно. Выход энергии в анаэробных процессах невелик, а количество энергии, необходимое микроорганизмам для синтеза клеточного вещества, такое же, как у аэробных форм. Поэтому у постоянных обитателей метантенка энергетический обмен преобладает над конструктивным.
Скорость реакций в метантенке может быть увеличена путем повышения температуры. Различают мезофильное сбраживание, при Т=30—33 °С и термофильное, при Т=50—55 °С. В последнем случае скорость распада органически веществ увеличивается, сокращается время сбраживания осадка и возрастает доза суточной загрузки в метантенк. Физиологические группы бактерий одинаковы в мезофильном и термофильном процессе, но среди терхмофилов больше спороносных форм. Большое преимущество термофильного сбраживания—то, что при Т=50—55 °С погибают яйца гельминтов, бактерии кишечной группы и другие патогенные неспоровые бактерии. Недостаток—трудность обезвоживания осадка.
Многие продукты анаэробного распада могут быть утилизированы. Выделяющийся в процессе метанового брожения метан собирается в газгольдеры и используется на отопление очистной станции, в том числе на поддержание требуемой температуры в метантенке. Сброженный осадок после подсушивания на иловых площадках или механического обезвоживания может быть использован в качестве удобрения на сельскохозяйственных полях. В Институте биохимии им. Баха разработан способ получения в процессе метанового брожения кормового витамина B12 при сбраживании отходов, богатых углеводами.