

Превращение и деструкция органического вещества
Биомасса автотрофных организмов—водорослей, высших водных растений и бактерий—служит источником питания для гетеротрофных организмов: бактерий, грибов, зоопланктона и нектона. Среди консументов выделяется особая группа организмов—редуцентов, которые минерализуют мертвое органическое вещество, тем самым обеспечивая продуцентов необходимыми им элементами минерального питания. К редуцентам относятся различные гетеротрофные микроорганизмы.
Бактерии и грибы разлагают отмершие водоросли (как и другие организмы), воздействуя на них экзоферментами. При этом часть сложных органических веществ минерализуется, а часть превращается в более простые соединения и в таком виде усваивается микроорганизмами. В клетках последних они подвергаются трансформации и входят в состав тел микроорганизмов в виде белков или запасных питательных веществ. Таким образом, микроорганизмы участвуют в деструкции органического вещества продуцентов и синтезируют органическое вещество собственного тела. Они являются первичными консументами. К первичным консументам относятся также все животные организмы, питающиеся растительной пищей, в том числе и растительноядные рыбы. Органическое вещество, образующее тело фитофагов, представляет вторичную продукцию. В дальнейшем это органическое вещество может еще раз трансформироваться, образуя третичную продукцию и т. д.
Для характеристики порядка превращения органических веществ существует понятие «трофический уровень». Продуценты образуют первый трофический уровень, фитофаги—второй, организмы, питающиеся фитофагами,— третий, и так далее. Переход органического вещества и заключенной в нем энергии с одного трофического уровня на другой осуществляется через пищевые цепи, в которых каждое предыдущее звено служит пищей последующему.
С переходом от предыдущего трофического уровня к последующему число и биомасса особей обычно снижаются, образуя так называемые пирамиды чисел или пирамиды биомасс. Мелкие организмы (протококковые водоросли, бактерии и др.) вследствие большой скорости размножения при одной и той же биомассе часто продуцируют больше органического вещества, чем крупные. При этом может создаваться такая ситуация, что биомасса организмов последующего трофического уровня будет больше предыдущей. В связи с этим необходимо хорошо отличать понятия «биомасса» и «продукция». Если биомасса—это суммарная масса всех имеющихся в наличии организмов данного трофического уровня, то продукция—это биомасса, образовавшаяся на данном трофическом уровне за определенный промежуток времени (в том числе и та биомасса, которая уже потреблена, отмерла или другим способом выведена из системы).
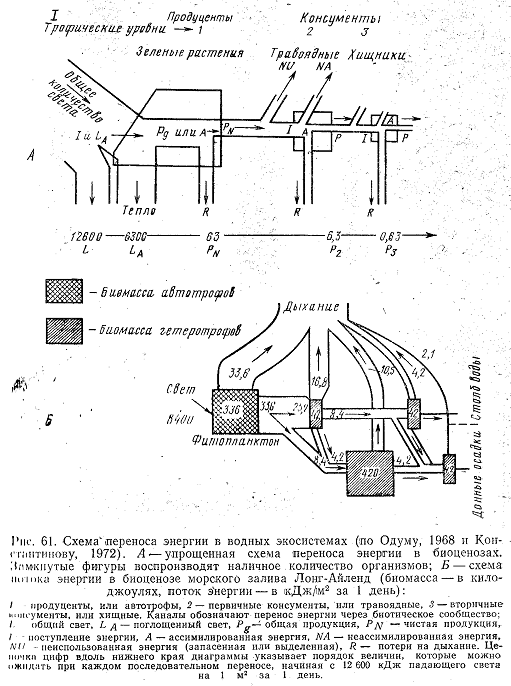
В отличие от биомассы энергия, заключенная в органическом веществе, при переходе с одного трофического уровня на другой непременно снижается. По этой причине Ю. Одумом, Г. Г. Винбергом и другими авторами предложен способ оценки продукции различных трофических уровней в эквивалентных единицах энергии. Пирамиды биомасс могут иметь основание более узкое, чем середину. Это наблюдается, например, в океане, где биомасса фитопланктона часто меньше биомассы организмов, питающихся фитопланктоном. В пирамидах энергии каждая следующая ступень всегда меньше предыдущей (рис. 61).
На любом трофическом уровне потребляемая организмом пища не усваивается полностью, остается некоторый остаток. Усвоенная часть пищи частично расходуется на прирост, а частично окисляется с выделением энергии. Та часть пищи, которая пошла на прирост, сохранила заключенное в ней количество энергии, и эта энергия может быть использована организмами следующего трофического уровня. Часть энергии, израсходованная в процессах жизнедеятельности организма, рассеивается в пространстве и исключается из дальнейшего превращения органического вещества. Однако вследствие того, что при рассеивании
энергии происходила и минерализация органического вещества, параллельно с освобождением энергии в воду поступали углекислота и биогенные элементы, которые способствовали образованию первичной продукции и, следовательно, новому накоплению энергии.
Сумма энергии, заключенной в приросте биомассы, потраченной в процессах обмена и оставшейся в неусвоенной части пищи, должна быть равна количеству энергии, первоначально содержавшемуся в пище, т. е.
![]()
где Р—пища (рацион), П—прирост биомассы, Т—траты на обмен, Н—неусвоенная пища. Все показатели выражены в одинаковых величинах, например, в единицах энергии или в количестве кислорода.
Эффективность использования пищи характеризуется безразмерными коэффициентами: K1—коэффициент использования потребленной пищи на рост, К2—коэффициент использования усвоенной пищи на рост.
![]()
Естественно, что коэффициенты K1 и К2 не могут быть равны единице, так как прирост тела без затраты энергии невозможен, но они могут быть равны нулю в том случае, когда прирост биомассы отсутствует. Чем выше коэффициент, тем большая часть рациона расходуется на прирост.
Коэффициенты K1 и К2 могут характеризовать превращение вещества не только организмами одного трофического уровня, но и системы, состоящей из двух и более трофических уровней. Например, сапрофитные бактерии, минерализующие органическое вещество водорослей, имеют K1 до 0,6. Это означает, что организмы, питающиеся бактериями, при полном потреблении продукции бактерий будут иметь рацион Р2 = 0,6 P1. Если допустить, что их коэффициент использования пищи на рост K1 тоже равен 0,6, хотя обычно он меньше, то они образуют продукции только 0,6х0,6=0,36 P1. В общем виде прирост системы, состоящей из двух трофических уровней, составит:
![]()
Так как K1 всегда меньше единицы, то чем больше трофических уровней в экосистеме, тем меньше конечный прирост биомассы в ней.
В природе практически не может быть полного потребления всей биомассы, образовавшейся на каком-либо трофическом уровне. Часть организмов отмирает естественным путем, другая по каким-то причинам выводится из системы и в результате только незначительная часть продукции используется организмами следующего трофического уровня. По мнению Я. Я. Цееба (1973), при каждом переходе органического вещества на последующие трофические уровни аккумулированная фитопланктоном энергия снижается примерно в 10 раз.
Количество трофических уровней в пелагиали чаще всего 3—4, а в бентосе 2—3. Например:

Биомасса хищных рыб составит незначительную часть от первичной продукции. Если же в водоеме живут только растительноядные рыбы, относящиеся ко второму трофическому уровню, то конечная биомасса будет значительно выше.
В последние годы предпринимаются успешные попытки построения математических моделей для описания процессов, протекающих в водоемах (Г. Г. Винберг и С. Н. Анисимов, 1966; В. В. Меншуткин, 1970; А. А. Умнов, 1972). Цель такого моделирования заключается в количественном изучении круговорота веществ и в установлении определяющих факторов, влияющих на продуктивность водоемов.