

Глава XIII. Превращение соединений металлов микроорганизмами
Многие микроорганизмы участвуют в превращении соединений металлов. Для одних это способ получения энергии, для других—побочная реакция. По направленности воздействия на соединения металлов можно выделить следующие группы бактерий: 1) окисляющие закисное железо с образованием гидрата окиси железа (железобактерии); 2) разлагающие органические комплексы железа и марганца; 3) окисляющие сульфиды металлов и способствующие выщелачиванию металлов из руд. Деление это весьма условии, так как у некоторых представителей первой группы образование Fe(ОН)3 наблюдается только в определенных условиях, а другие бактерии по осуществляемым реакциям могут быть отнесены одновременно к двум группам.
Окисление соединений записного железа
Железо присутствует в воде водоемов чаще всего в виде растворимых закисных солей карбонатов Fе(НСО3)2 и FeCO3. В присутствии кислорода эти соли переходят в гидрат окиси железа:
![]()
Процесс протекает с выделением энергии. Образовавшаяся гидроокись железа выпадает в осадок в виде бурых (охристых) хлопьев. Эта реакция может проходить и чисто химическим путем, и под влиянием микроорганизмов.
Среди бактерий, окисляющих закисное железо с образованием гидрата окиси железа Fe(OH)3, известны нитчатые и одноклеточные формы. Большинство из них способны отлагать не только окись железа, по и окись марганца, если марганец присутствует и воде.
НИТЧАТЫЕ ЖЕЛЕЗОБАКТЕРИИ
Нитчатые
железобактерии впервые были выделены
и изучены С Н. Виноградским, который
отнес их к автотрофным организмам,
получающим энергию в процессе окисления
закисного железа в окисное. Нитчатые
железобактерии чрезвычайно широко
распространены в природе. В воде лесных
ручьев, в болотах и других водоемах
часто можно видеть бурые хлопья, большей
частью вытянутые по течению. П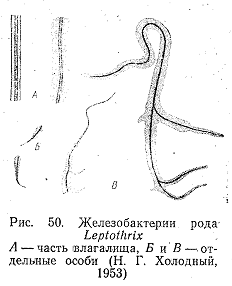 ри
микроскопическом исследовании видно,
что эти хлопья представляют собой
длинные слизистые трубки, внутри которых
иногда находятся бактериальные клетки.
Слизистые чехлы дают реакцию на окисное
железо и представляют продукт
жизнедеятельности видов рода Leptothrix.
Наиболее распространенный вид данного
рода—L.
ochracea
(рис.
50). Окисляя закисное железо, он выделяет
гидроокись железа, которая откладывается
на поверхности клеток. При этом образуется
типичный слизистый чехол, который
препятствует сообщению клеток с внешней
средой. Когда чехол становится достаточно
плотным, клетки покидают его (рис. 51).
Освободившиеся клетки имеют жгутики и
потому подвижны. Покинув старый чехол,
они приступают к выработке нового. Таким
образом, в водоеме накапливаются пустые
чехлы, содержащие окисное железо. Чехол,
или влагалище, L.
ochracea имеет
форму цилиндра, диаметр которого
постоянен на всем протяжении. Внутренний
диаметр трубки 1 мкм, наружный—2—3 мкм.
ри
микроскопическом исследовании видно,
что эти хлопья представляют собой
длинные слизистые трубки, внутри которых
иногда находятся бактериальные клетки.
Слизистые чехлы дают реакцию на окисное
железо и представляют продукт
жизнедеятельности видов рода Leptothrix.
Наиболее распространенный вид данного
рода—L.
ochracea
(рис.
50). Окисляя закисное железо, он выделяет
гидроокись железа, которая откладывается
на поверхности клеток. При этом образуется
типичный слизистый чехол, который
препятствует сообщению клеток с внешней
средой. Когда чехол становится достаточно
плотным, клетки покидают его (рис. 51).
Освободившиеся клетки имеют жгутики и
потому подвижны. Покинув старый чехол,
они приступают к выработке нового. Таким
образом, в водоеме накапливаются пустые
чехлы, содержащие окисное железо. Чехол,
или влагалище, L.
ochracea имеет
форму цилиндра, диаметр которого
постоянен на всем протяжении. Внутренний
диаметр трубки 1 мкм, наружный—2—3 мкм.
Нити L. ochracea никогда не ветвятся и не прикрепляются к субстрату. Развиваются они обычно в слабоминерализованной воде, концентрация железа 0,1 мг/л обеспечивает обильный рост клеток. Физиология L. ochracea изучена недостаточно, в частности не установлен определенно тип питания микроорганизма. Известно, что ему требуется закисное железо, но, возможно, он нуждается и в органических веществах.

Другой вид—L. crassa отличается от L. ochracea образом жизни и морфологией. L. crassa может свободно плавать, но способен вести и прикрепленный образ жизни. Нити его обычно различаются по толщине. Прикрепленный конец влагалища широкий, диаметр его достигает 10—15 мкм. Другой конец постепенно истончается, и конечные растущие клетки не имеют чехла (см. рис. 50). В состав чехла, помимо гидроокиси железа, входит органические вещества, По типу питания L. crassa относится к литогетеротрофам.
Микроорганизмы рода Leptofhrix играют важную роль п процессах образования болотных железных руд.
Ж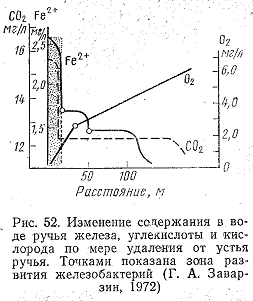 елезобактерии
создают серьезные помехи в водоснабжении.
Почвенные и подземные йоды часто содержат
железо в закисной форме и углекислоту.
При выходе на дневную поверхность воды
теряют углекислоту и насыщаются
кислородом. При этом закисное железо
переходит в окисное. Рост железобактерий
в этих условиях может оказаться очень
обильным. На рис. 52 показано, как меняется
в воде источника содержание железа,
углекислоты и кислорода по мере удаления
от устья ручья, и выделена область
развития железобактерий и ручье.
Максимальное развитие L.
ochracea
наблюдается в зоне, где концентрации
Fe2+
превышает 2,5 мг/л, CO2—не
менее 12,5 мг/л, а кислорода меньше 2 мг/л.
елезобактерии
создают серьезные помехи в водоснабжении.
Почвенные и подземные йоды часто содержат
железо в закисной форме и углекислоту.
При выходе на дневную поверхность воды
теряют углекислоту и насыщаются
кислородом. При этом закисное железо
переходит в окисное. Рост железобактерий
в этих условиях может оказаться очень
обильным. На рис. 52 показано, как меняется
в воде источника содержание железа,
углекислоты и кислорода по мере удаления
от устья ручья, и выделена область
развития железобактерий и ручье.
Максимальное развитие L.
ochracea
наблюдается в зоне, где концентрации
Fe2+
превышает 2,5 мг/л, CO2—не
менее 12,5 мг/л, а кислорода меньше 2 мг/л.
П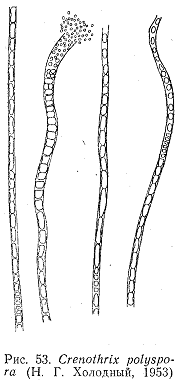 ри
поступлении таких вод в трубопроводы
в них также развиваются железобактерии.
Иногда при их интенсивном размножении
сужается сечение трубопровода. Для
предотвращения этого нежелательного
явления необходимо удалить соли железа
из воды до поступления ее в трубопровод.
Это достигается аэрацией воды с
последующим отстаиванием. Помехи в
водоснабжении может создавать и другая
нитчатая железобактерия—Crenothrix.
Для нее характерны коническая форма
влагалища и прикрепление к субстрату
узким концом. Железо отлагается только
у основания. Длина нитей Crenolhrix
достигает 3 мм, ширина у основания
составляет 1,5—5,0 мкм, а у верхнего конца
6—9 мкм. Клетки Crenothrix
размножаются не только поперечным, но
и продольным делением, поэтому у открытого
конца влагалища образуется несколько
параллельных рядов (рис. 53). Клетки
выходят из влагалища, прикрепляются к
субстрату и дают начало новым нитям.
Кроме того, Crenothrix
способен размножаться короткими
обрывками нитей. И конце прошлого века
в ряде городов Западной Европы (Амстердам,
Берлин, Дрезден) массовое развитие
Crenothrix
вывело из строя городские водопроводы.
ри
поступлении таких вод в трубопроводы
в них также развиваются железобактерии.
Иногда при их интенсивном размножении
сужается сечение трубопровода. Для
предотвращения этого нежелательного
явления необходимо удалить соли железа
из воды до поступления ее в трубопровод.
Это достигается аэрацией воды с
последующим отстаиванием. Помехи в
водоснабжении может создавать и другая
нитчатая железобактерия—Crenothrix.
Для нее характерны коническая форма
влагалища и прикрепление к субстрату
узким концом. Железо отлагается только
у основания. Длина нитей Crenolhrix
достигает 3 мм, ширина у основания
составляет 1,5—5,0 мкм, а у верхнего конца
6—9 мкм. Клетки Crenothrix
размножаются не только поперечным, но
и продольным делением, поэтому у открытого
конца влагалища образуется несколько
параллельных рядов (рис. 53). Клетки
выходят из влагалища, прикрепляются к
субстрату и дают начало новым нитям.
Кроме того, Crenothrix
способен размножаться короткими
обрывками нитей. И конце прошлого века
в ряде городов Западной Европы (Амстердам,
Берлин, Дрезден) массовое развитие
Crenothrix
вывело из строя городские водопроводы.
Нитчатая бактерия Sphaerotilus natans по многим признакам очень близка к железобактериям, поэтому ее обычно рассматривают вместе с родом Leptothrix. Sphaerotilus образует длинные нити, состоящие из цепочек клеток, окруженных общим влагалищем. В отличие от Leptothrix влагалище Sphaerotilus состоит из полисахаридов. Клетки размножаются внутри влагалища поперечным делением. Они имеют полярные жгутики, могут покидать влагалище, прикрепляться к субстрату и прорастать. Очень часто бактерии прикрепляются к тому влагалищу, которое покинули. Это создает впечатление ложного ветвления нитей. Типичный вид рода Sphaerotilus—Sph. natans. Он относится к гетеротрофам и растет в сильно загрязненных и сточных водах. Обладает весьма активным обменом и дает вспышки массового развития. Влагалище Sph. natans обычно не содержит солей железа, по экспериментально показано, что при культивировании па специальных средах в нем возможно отложение солей железа. Марганец в чехлах Sph. natans не откладывается.
Sph. natans дает мощные разрастания в водоемах и водопроводных трубах. Бороться с этой бактерией чрезвычайно трудно. На очистных сооружениях развитие Sph. natans приводит к так называемому «вспуханию» активного ила. Упругие нити бактерий пронизывают активный ил и препятствуют его осаждению. Все другие организмы из аэротенков вытесняются. Меры борьбы с развитием Sph. natans—соблюдение технологического режима, усиленная аэрация, подщелачивание среды в аэротенках до рН 9. Нити Sphaerotilus при этом распадаются на отдельные клетки. В результате подщелачивания гибнет большая часть биоценоза аэротенка, но в течение нескольких суток активность ила восстанавливается.
ОДНОКЛЕТОЧНЫЕ ЖЕЛЕЗОБАКТЕРИИ
Типичный представитель одноклеточных железобактерий—Gallionella. Эта бактерия в чистой культуре не получена и потому изучена недостаточно. Бактерии этого рода представляют собой слегка изогнутые бобовидные клетки. От вогнутой стороны клетки отходят длинные переплетенные «стебельки». Стебельки хорошо различимы в микроскоп, и их форма служит систематическим признаком. За каждый час Gallionella выделяет стебелек, в 30 раз превышающий толщину клетки. Общая длина стебелька может достигать 1000 мкм. В состав стебелька входят железо (вероятно, и аморфном состоянии) и органическое вещество. По сравнению со с стебельком сама клетка очень мала, ее размеры 0,5—0,6х1,2—1,5 мкм, и по этой причине клетка Gallionella долго оставалась незамеченной исследователями.
Gallionella получает энергию в процессах окисления закисного железа. Она способна расти на минеральной среде без органических веществ, следовательно, по типу обмена этот микроорганизм скорее всего относится к литоавтотрофам. Выход энергии у Gallionella настолько мал, что для синтеза 1 г клеток бактерии необходимо выделить 500 г гидроокиси железа. Более 90 % от сухой массы организма приходится на окисное железо.
Gallionella развивается в железистых водах при значениях рН, близких к нейтральному, и при пониженной концентрации кислорода. Она способна расти в холодной воде и даже под снегом. Являясь активным участником процессов отложения окисного железа, они и отличие от многих других железобактерий никогда не отлагает соединений марганца. Вследствие активного продуцирования стебельков Gallionella может вызывать закупорку водопроводных труб и создавать серьезные помехи в водоснабжении.