

Скорость роста и физиологическая активность
Физиологическая активность измеряется количеством питательных веществ, потребляемых единицей микробной биомассы за единицу времени, или количеством образующихся продуктов:
![]()
где q—физиологическая активность; а—трофический коэффициент, показывающий затраты питательных веществ на образование единицы биомассы; b—коэффициент основного обмена, показывающий расход питательных веществ на поддержание жизни единицы биомассы в течение 1 ч.
Вещества, потребляемые микроорганизмами, расходуются на поддержание жизненных процессов в клетке. Это так называемый основной обмен.
Коэффициенты
а и b
для определенной культуры и условий—величины
постоянные, и таким образом физиологическая
активность зависит от удельной скорости
роста. При интенсивном росте культуры
aμ>b,
физиологическая активность высокая.
При μ=0 первый член уравнения также равен
нулю, и физиологическая активность
становится равной основному обмену.
Следовательно, высокой физиологической
активностью обладает только быстрорастущая
культура. Это положение било подтверждена
в экспериментах Н. Д. Иерусалимского с
сотрудниками на примере Azotobacter vinelandii,
выращиваемого в проточных условиях
(рис. 33).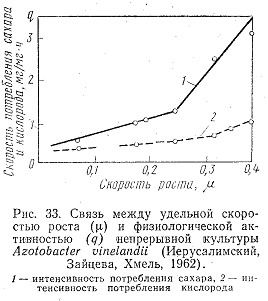
При низкой скорости роста потребляемые продукты больше расходуются на основной обмен, при ускорении роста большая часть потребляемого сахара идет на биосинтез. Таким образом, низкая скорость роста сопровождается более глубоким распадом усвоенных продуктов питания, высокая же скорость приводит к превращению органических веществ из одной формы в другую, в тело микроорганизмов.
Прирост биомассы по отношению к потребленному субстрату называют экономическим коэффициентом (у). Он представляет собой частное от деления скорости роста на физиологическую активность микроорганизмов:
![]()
При снижении скорости роста знаменатель возрастает, и следовательно, экономический коэффициент снижается. При увеличении μ экономический коэффициент увеличивается.
Естественно, что для микробиологического синтеза, например для получения кормовых дрожжей, желательно вести процесс в условиях, обеспечивающих высокую скорость роста культуры. Для глубокой очистки сточных вод, когда требуется возможно более полно окислить содержащиеся в них органические вещества, приходится вести процесс при относительно невысокой скорости роста.
Глава VI. Влияние внешних факторов на микроорганизмы Влияние температуры
Действие температуры испытывают все живые организмы, в том числе и микроскопические. По отношению к температуре микроорганизмы делятся на мезофилы, психрофилы и термофилы.
Мезофилы (мезо—средний) лучше всего приспособлены к существованию в интервале средних температур. Они могут нормально размножаться в пределах от 20 до 40 °С. В этой группе, как и в двух других, есть организмы, развивающиеся в более широком или более узком диапазоне температур, и указанный интервал нельзя считать строго ограниченным. К мезофилам относятся большая часть микроорганизмов.
Психрофилы (психрос—холод) приспособлены к существованию при пониженных температурах. Они нормально живут и размножаются при температуре от —8 до 10 °С. Большинство психрофилов способно расти и при температурах, характерных для мезофилов, поэтому их называют факультативными, т. е. необязательными психрофилами. В отличие от факультативных облигатные (т. е. обязательные) психрофилы погибают при температурах, близких к 30 °С. К психрофилам относятся некоторые почвенные и морские бактерии, а также виды, патогенные для морских животных и растений. Некоторые психрофилы вызывают порчу продуктов, хранящихся при пониженных температурах.
Термофилы (термо—тепло) способны нормально существовать при температурах от 50 до 75 °С. Они обнаруживаются в горячих источниках, в молоке, почве, навозе. Они не погибают при пастеризации молочных продуктов, некоторые из них выдерживают 10-минутное нагревание при температурах 80—90 °С. Следует отличать выживаемость при какой-либо температуре и оптимальное значение температуры для жизнедеятельности микроорганизмов. Оптимум обычно лежит ближе к верхней границе температурного диапазона. При более высоких температурах процессы жизнедеятельности активируются, организмы быстрее растут и размножаются. Это связано с активацией работы ферментов при повышении температуры. Превышение границы температурного интервала, как правило, приводит к разрушению ферментов и к гибели вегетативных клеток. При температуре 70 °С большинство вегетативных клеток мезофилов погибает в течение 1—5 мин, при температуре 63 °С—в течение 30 мин. Термофилы при такой температуре способны расти и размножаться. Причина повышенной термостабильности термофилов заключается, во-первых, в качественном и количественном своеобразии отдельных компонентов клеток (мембран, рибосом и особенно оболочек) и, во-вторых, в более, быстром синтезе клеточных элементов. Благодаря последнему менее стабильные компоненты клетки, например РНК, быстро заменяются при разрушении. Адаптация микроорганизмов к повышенным температурам очень затруднена. Споры более устойчивы к нагреванию, чем вегетативные клетки, причем споры термофилов устойчивее, чем споры мезофилов. При кипячении все вегетативные клетки погибают за несколько секунд, а споры многих видов микроорганизмов выдерживают кипячение в течение нескольких часов. Понижение температуры переносится микроорганизмами гораздо лучше, чем нагревание. У многих видов вегетативные клетки способны выдерживать однократное замораживание. Причину гибели клеток при замораживании многие исследователи видят в разрушающем действии кристаллов льда, образующихся в клетках. В случае быстрого замораживания (1—10 с), когда вместо кристаллов в клетках образуется стекловидная ледяная масса, процент погибших клеток значительно снижается.