

Получение энергии литотрофами
Литотрофы получают энергию в процессах фото- и, хемосинтеза. В процессе фотосинтеза источником энергии служит солнечный свет, донором электронов—водород неорганических соединений, воды или сероводорода. Фотосинтезирующие литотрофные организмы называют фотолитотрофами. Уравнение фотосинтеза в общем виде может быть представлено формулой:
![]()
Где А—донор водорода.
У фотосинтезирующих пурпурных серобактерий в качестве донора водорода выступает сероводород, у растений—вода.
Это уравнение не отражает всей сложности процесса фотосинтеза. В настоящее время фотосинтез растений хорошо изучен. Он осуществляется с помощью зеленого пигмента хлорофилла. Строение хлорофилла имеет много общего со строением гема, являющегося активной группой цитохромов, каталазы и других ферментов, но в отличие от них в состав хлорофилла вместо железа входит магний.
Строение молекулы хлорофилла благодаря чередованию одинарных и двойных связей, расположенных по кольцу, обусловливает его активность. В молекуле хлорофилла есть электроны, не связанные с определенным атомом, а принадлежащие целиком молекуле. Такие электроны особенно легко приходят в возбужденное состояние под действием световых лучей. Как известно, свет состоит из элементарных частиц—фотонов и вместе с тем имеет волновую природу. Энергия каждого фотона обратно пропорциональна длине его волны, т. е. наибольшей энергией обладают фотоны коротковолновой части спектра (рентгеновские и ультрафиолетовые лучи), а наименьшей—инфракрасные лучи и радиоволны. Получая энергию, электрон возбуждается и переходит на другую орбиту, характеризующуюся более высоким энергетическим уровнем.
Заняв высокий энергетический уровень, электрон может перейти на более низкий, т. е. вернуться на первоначальную орбиту. Если это происходит, то энергия выделяется в виде света, и наблюдается люминесценция. При переходе электрона с орбиты на орбиту часть энергии рассеивается, и поэтому наблюдаемое свечение (люминесценция) имеет большую длину волны, чем поглощенные лучи.
В клетках зеленых растений электрон, выбитый квантом света со своей орбиты, вместе с воспринятой им энергией улавливается цепью ферментов. В передаче электронов участвуют флавоноиды, цитохромы, железосодержащий белок ферредоксин и многие ферменты. В результате передачи электрона по цепи он снова возвращается к хлорофиллу, который, потеряв электрон, приобрел положительный заряд и тем самым стал возможным акцептором электронов.
По пути электрона к хлорофиллу часть его энергии дважды расходуется на присоединение фосфорной кислоты к АДФ. В результате образуются две молекулы АТФ. Этот процесс носит название фотосинтетического фосфорилирования (рис. 26).
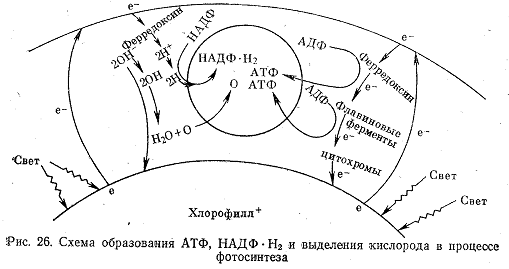
Вследствие диссоциации воды в клетке всегда содержится некоторое количество ионов Н+ и ОН-. Часть возбужденных электронов захватывается ионами водорода, которые при этом превращаются в атомы. Свободные атомы водорода в химическом отношении весьма активны, они присоединяются к первичной дегидрогеназе НАДФ (с. 49). НАДФ при этом переходит в восстановленную форму НАДФ·Н2. На этот процесс также расходуется часть энергии фотонов, воспринятой электроном. Таким образом, в результате возбуждения электронов фотонами в клетке образуется АТФ и НАДФ·Н2. Оставшиеся без противоиона ионы ОН- отдают свой электрон (есть основания полагать, что за счет этого электрона происходит восстановление хлорофилла) и превращаются в радикал ОН. Радикалы реагируют между собой:
![]()
Следовательно, выделяющийся в процессе фотосинтеза кислород образуется из воды, а не из углекислоты. Это было подтверждено экспериментально в опытах с мечеными атомами 18О.
Процессы образования АТФ, НАДФ·Н2 и выделения кислорода происходят на свету. Усвоение клеткой углекислого газа и синтез из него органических веществ может протекать и в темноте. Это показали Арнон и его сотрудники. Они облучали изолированные хлоропласты светом в отсутствие углекислого газа, но в присутствии АДФ и НАДФ. При этом в хлоропластах накапливались АТФ и НАДФ·Н2 и выделялся кислород. Затем из хлоропластов были выделены ферменты, АТФ, НАДФ·Н2 и в темноте, в присутствии СО2, наблюдался синтез углеводов. В других опытах к фотосинтезирующим ферментам были добавлены АТФ и НАДФ·Н2, выделенные из животных клеток. При этом также наблюдался синтез углеводов. Таким образом, ученым удалось показать единство процессов биосинтеза у растений и животных.
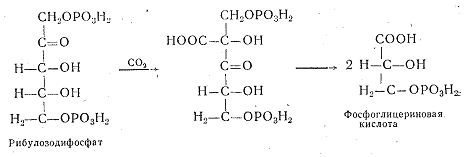
Механизм ассимиляции молекулы СО2 был открыт М. Кальвином в экспериментах с радиоактивным углеродом 14С. М. Кальвин доказал, что в процессе фотосинтеза СО2 присоединяется к органическому соединению рибулозодифосфату, который при этом превращается в две молекулы фосфоглицериновой кислоты.
Процесс носит циклический характер (рис. 27). В результате ряда превращений из фосфоглицериновой кислоты образуется гексоза фруктозодифосфат, которая используется на построение крахмала и образование рибулозодифосфата. Фиксация углекислоты протекает с поглощением энергии, накопленной в световую стадию. Энергия черпается из АТФ, которая при этом переходит в АДФ. Водород НАДФ·Н2 расходуется на восстановление фосфоглицериновой кислоты.
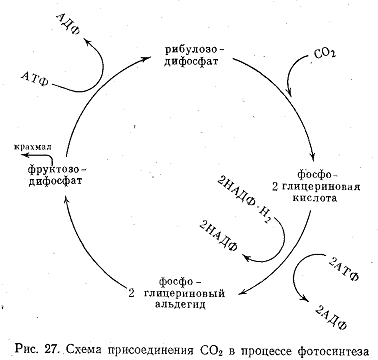
В процессе ассимиляции углекислоты на восстановление одной молекулы СО2 расходуется 3 молекулы АТФ и 4 атома водорода. При этом в среднем затрачивается 5—6 квантов энергии. На площади 1 м2 зеленых листьев в течение 1 ч синтезируется примерно 1 г органического вещества и выделяется около 1,7 г кислорода. Фотосинтез свойствен и некоторым бактериям.
В процессе хемосинтеза получают энергию лишь некоторые группы бактерий: нитрифицирующие, сероокисляющие, водородные и железобактерии. Выделившуюся при хемосинтезе энергию микроорганизмы используют в процессах конструктивного обмена. Процессы обмена веществ у хемосинтезирующих бактерий имеют много общего с процессами у фотосинтезирующих. Ассимиляция углекислоты идет у них, по-видимому, тоже путем присоединения СО2 к молекуле рибулозодифосфата, а выделившаяся энергия запасается в макроэргических связях АТФ.
Энергетически процесс хемосинтеза малоэффективен. По этой причине хемосинтезирующим бактериям для обеспечения энергией реакции присоединения одной молекулы углекислоты приходится окислять несколько десятков молекул субстрата.
Процессы получения энергии различными видами литотрофных микроорганизмов могут протекать как в присутствии кислорода (аэробные условия), так и в его отсутствие (анаэробные условия). В аэробных условиях осуществляют процессы окисления железобактерии, нитрифицирующие, водородные и большинство тионовых бактерий. Акцептором электронов у них выступает кислород.
В отсутствие молекулярного кислорода акцепторами электронов могут быть неорганические соединения, такие как углекислота. Процесс, в котором окисление осуществляется за счет связанного кислорода, носит название анаэробного дыхания. Примером анаэробного дыхания может служить окисление водорода метановыми бактериями.
![]()
При анаэробном дыхании, так же как при других окислительных процессах, имеет место окислительное фосфорилирование, т. е. образование АТФ. Анаэробное дыхание свойственно и литотрофам, и органотрофам.