Регуляция синтеза ферментов
Несмотря на малые размеры клетки, в ней одновременно присутствует несколько тысяч различных химических соединений. Количество одних соединений поддерживается на постоянном уровне, других—варьирует в широких пределах. АТФ, например, обычно составляет 0,04 % от массы клетки. В период интенсивного биосинтеза, когда АТФ быстро расходуется, активно протекают и окислительные процессы, позволяющие клетке компенсировать затраченную энергию и восполнить запас АТФ.
Число белков, одновременно присутствующих в клетке, может достигать нескольких сотен и даже тысяч. В сбалансированном состоянии концентрация каждого белка составляет от нескольких сотых до нескольких десятых процента от суммы всех белков. Если же в клетке возникает недостаток какого-либо жизненно важного соединения, в ней начинается усиленный синтез ферментов, способствующих образованию именно этого соединения. Так, при недостатке в клетке фосфорной кислоты (необходимой для образования АТФ) начинается усиленный синтез фосфатазы, вызывающий омыление эфиров фосфорной кислоты и выделение фосфатов в свободном состоянии. Количество фосфатазы при этом может увеличиться в 50—100 раз и достигнуть 5 % от всех белков клетки. Весьма значительно изменяется содержание окислительно-восстановительных ферментов, дегидрогеназ, зависящее от интенсивности окислительно-восстановительных процессов в клетке. Чем активнее клетка окисляет субстрат, тем выше в ней концентрация дегидрогеназ. (Благодаря этому оказалось возможным характеризовать степень активности ила на очистных сооружениях по содержанию в нем дегидрогеназ. Чем интенсивнее ил перерабатывает загрязнения, тем выше его дегидрогеназная активность).
Все протекающие в клетке химические реакции хорошо сбалансированы друг с другом. Это свидетельствует о существовании в клетке ряда систем автоматического регулирования, управляющих кинетикой обменных процессов. Гипотезу регулирования синтеза ферментов в клетке предложили французские ученые Ф. Жакоб и Ж. Моно. Согласно этой гипотезе, все гены, ответственные за синтез какого-либо соединения, располагаются в молекуле ДНК последовательно один за другим, в порядке действия ферментов. Так как эти гены определяют структуру фермента, их называют структурными (рис. 22). Кроме них существует еще и ген-оператор, «включающий» и «выключающий» структурные гены. Ген-оператор ответственен за одновременную активность
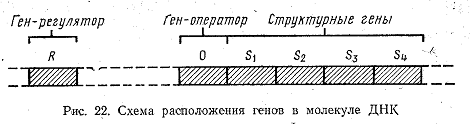
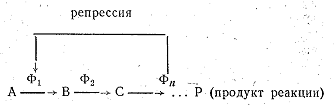
структурных генов. Он располагается на одном конце цепи структурных генов и непосредственно воздействует на них. Не исключено, что ген-оператор не самостоятельный ген, а входит в первый структурный ген. Структурные гены и ген-оператор вместе составляют оперон. Установлено, что сигнал на включение и выключение ген-оператор получает извне, от так называемого гена-регулятора, который располагается в той же молекуле ДНК, но на некотором расстоянии от оперона. Ген-регулятор действует на оперон посредством вещества, выделяемого им в цитоплазму. Вещество, влияющее на оперон и подавляющее синтез ферментов, называется репрессором. Оно, как многие ферменты, состоит из двух компонентов: первый выделяется геном-регулятором, а второй является низкомолекулярным соединением. По аналогии с ферментами компоненты репрессора называются апорепрессором (выделяется геном-регулятором) и корепрессором. В отдельности каждый из них неактивен, но, соединяясь вместе, они образуют активный репрессор. В качестве корепрессора обычно выступает конечный продукт реакции, осуществляемой с помощью данного оперона. Когда в клетке накапливается значительное количество синтезируемого продукта, он соединяется с апорепрессором, выделяемым геном-регулятором. В результате образуется репрессор, который воздействует на ген-оператор и связывает его. При этом ген-оператор оказывается выключенным, он не подает сигнала структурным генам и синтез данного вещества прекращается (рис. 23). Если конечный продукт
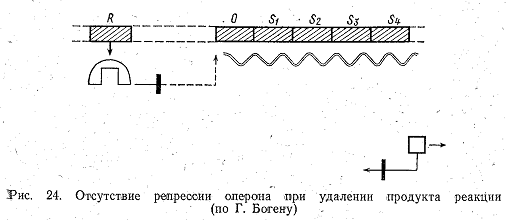
реакции используется клеткой и не накапливается, репрессор не образуется, оперон остается включенным, и синтез продолжается (рис. 24). Система торможения синтеза конечным продуктом реакции позволяет клетке работать экономично и синтезировать только те соединения, которые ей необходимы, но она имеет один недостаток. Пока конечный продукт реакции будет связываться с апорепрессором, а репрессор выключать ген-оператор, синтез и-РНК на опероне будет продолжаться, и пройдет еще некоторое время, прежде чем распадутся все уже синтезированные молекулы и-РНК, ответственные за синтез данного продукта. Видимо, поэтому в клетке имеется и другой механизм, моментально прекращающий синтез продукта, необходимость в котором отпала. Он заключается в том, что продукт реакции непосредственно связывается с первым в цепи ферментов, катализирующих синтез, и блокирует его.
Эти два механизма регуляции синтеза взаимно дополняют друг друга.
Система регуляции расщепления каких-либо продуктов имеет сходный характер с регуляцией синтеза, но в этом случае низкомолекулярное соединение не выключает, а включает ген-оператор. Этот механизм удобно рассмотреть на примере расщепления сахара лактозы. Лактоза ферментом β-галактозидазой расщепляется на глюкозу и галактозу. Глюкоза—наиболее часто встречающийся питательный субстрат, и ферменты для ее расщепления: присутствуют в клетке постоянно. Это конститутивные ферменты. Лактоза используется реже, для ее расщепления требуются индукционные ферменты.
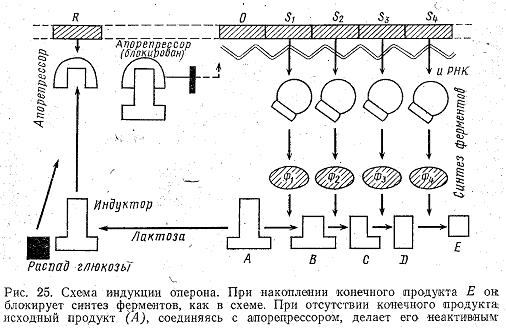
Синтез в клетке индукционных ферментов обычно подавлен, причем корепрессором для синтеза ферментов, расщепляющих лактозу, служит продукт расщепления глюкозы. Пока глюкоза присутствует в клетке и может расщепляться, клетке незачем затрачивать энергию на синтез индукционных ферментов. Когда же запас корепрессора в клетке истощается, в действие вступает еще один фактор, называемый индуктором. Индуктор способен связываться с апорепрессором, т. е. он конкурент корепрессору. При большом количестве корепрессора в клетке индуктор не имеет возможности связаться с апорепрессором, когда же корепрессор иссякает, индуктор связывается с апорепрессором и блокирует его. При этом ген-оператор, не испытывая репрессии гена-регулятора, оказывается «включенным», что и приводит к синтезу соответствующих индукционных ферментов (рис. 25). Индуктором: служит то самое вещество, для расщепления которого требуются индукционные ферменты, в приведенном примере—лактоза.
Из сказанного следует, что вещество, продуцируемое геном-регулятором, может при соединении с индуктором служить сигналом для синтеза белка, а при соединении с корепрессором—сигналом прекращения синтеза. В этом проявляется экономичность и высокая эффективность систем регулирования в клетке.