

7.Зарубежные исследователи, внесшие наибольший вклад в становлении физиологии как науки.
Исторический очерк. Первоначальные сведения из области Физиология были получены в глубокой древности на базе эмпирических наблюдений натуралистов и врачей и особенно анатомических вскрытий трупов животных и людей. На протяжении многие веков во взглядах на организм и его отправления господствовали идеи Гиппократа (5 в. до н. э.) и Аристотеля (4 в. до н. э.). Однако наиболее существенный прогресс Физиология был определён широким внедрением вивисекционных экспериментов, начало которых было положено ещё в Древнем Риме Галеном (2 в. до н. э.). В средние века накопление биологических знаний определялось запросами медицины. В эпоху Возрождения развитиюФизиология способствовал общий прогресс наук. Физиология как наука ведёт своё начало от работ английского врача У. Гарвея, который открытием кровообращения (1628) «... делает науку из физиологии (человека, а также животных)» (Энгельс Физиология, Диалектика природы, 1969, с. 158). Гарвеем были сформулированы представления о большом и малом кругах кровообращения и о сердце как двигателе крови в организме. Гарвей первый установил, что кровь по артериям течёт от сердца и по венам возвращается к нему. Основу для открытия кровообращения подготовили исследования анатомов А. Везалия, испанского учёного М.Сервета (1553), итальянского – Р. Коломбо (1551), Г. Фаллопия и др. Итальянский биолог М. Мальпиги, впервые (1661) описавший капилляры, доказал правильность представлений о кровообращении. Ведущим достижением Физиология, определившим её последующую материалистическую направленность, явилось открытие в 1-й половине 17 в. французским учёным Р. Декартом и позже (в 18 в.) чеш. врачом Й. Прохаской рефлекторного принципа, согласно которому всякая деятельность организма является отражением – рефлексом – внешних воздействий, осуществляющихся через центральную нервную систему. Декарт предполагал, что чувствительные нервы являются приводами, которые натягиваются при раздражении и открывают клапаны на поверхности мозга. Через эти клапаны выходят «животные духи», которые направляются к мышцам и вызывают их сокращение. Открытием рефлекса был нанесён первый сокрушит, удар церковно-идеалистическим представлениям о механизмах поведения живых существ. В дальнейшем «... рефлекторный принцип в руках Сеченова стал оружием культурной революции в шестидесятых годах прошлого столетия, а через 40 лет в руках Павлова он оказался мощным рычагом, повернувшим на 180° всю разработку проблемы психического» (Анохин П. К., От Декарта до Павлова, 1945, с. 3). В 18 в. в Физиология внедряются физические и химические методы исследования. Особенно активно применялись идеи и методы механики. Так, итальянский учёный Дж. А. Борелли ещё в конце 17 в. использует законы механики для объяснения движений животных, механизма дыхательных движений. Он же применил законы гидравлики к изучению движения крови в сосудах. Английский учёный С. Гейлс определил величину кровяного давления (1733). Французский учёный Р. Реомюр и итальянский натуралист Л. Спалланцани исследовали химизм пищеварения. Франц. учёный А. Лавуазье, исследовавший процессы окисления, пытался на основе химических закономерностей приблизиться к пониманию дыхания. Итальянский учёный Л. Гальвани открыл «животное электричество», т. е. биоэлектрические явления в организме. К 1-й половине 18 в. относится начало развития Физиология в России. В открытой в 1725 Петербургской АН была создана кафедра анатомии и Физиология Возглавлявшие её Д. Бернулли, Л. Эйлер, И. Вейтбрехт занимались вопросами биофизики движения крови. Важными для Физиология были исследования М. В. Ломоносова, придававшего большое значение химии в познании физиологических процессов. Ведущую роль в развитии Физиология в России сыграл медицинский факультет Московского университета, открытого в 1755. Преподавание основ Физиология вместе с анатомией и др. медицинскими специальностями было начато С. Г. Зыбелиным. Самостоятельная кафедра Физиология в университете, которую возглавили М. И. Скиадан и И. И. Вечь, была открыта в 1776. Первая диссертация поФизиология выполнена Физиология И. Барсук-Моисеевым и посвящена дыханию (1794). В 1798 была основана Петербургская медико-хирургическая академия (ныне Военно-медицинская академия им. С. М. Кирова), где в дальнейшем Физиология также получила значительное развитие. В 19 в. Физиология окончательно отделилась от анатомии. Определяющее значение для развития Физиология в это время имели достижения органической химии, открытие закона сохранения и превращения энергии, клеточного строения организма и создание теории эволюционного развития органического мира. В начале 19 в. считали, что химические соединения в живом организме принципиально отличны от неорганических веществ и не могут быть созданы вне организма. В 1828 нем. химик Физиология Вёлер синтезировал из неорганических веществ органическое соединение – мочевину и тем самым подорвал виталистические представления об особых свойствах химических соединений организма. Вскоре нем. учёный Ю. Либих, а затем и многие другие учёные синтезировали различные органические соединения, встречающиеся в организме, и изучили их структуру. Эти исследования положили начало анализу химических соединений, участвующих в построении организма и обмене веществ. Развернулись исследования обмена веществ и энергии в живых организмах. Были разработаны методы прямой и непрямой калориметрии, позволившие точно замерять количество энергии, заключённой в различных пищевых веществах, а также освобождаемой животными и человеком в покое и при работе (работы В. В. Пашутина, А. А. Лихачева в России, М. Рубнера в Германии, Физиология Бенедикта, У. Этуотера в США и др.); определены нормы питания (К. Фойти др.). Значительное развитие получила Физиология нервно-мышечной ткани. Этому способствовали разработанные методы электрического раздражения и механической графической регистрации физиологических процессов. Нем. учёный Э. Дюбуа-Реймон предложил санный индукционный аппарат, нем. физиолог К. Людвиг изобрёл (1847) кимограф, поплавковый манометр для регистрации кровяного давления, кровяные часы для регистрации скорости кровотока и пр. Французский учёный Э. Марей первый применил фотографию для изучения движений и изобрёл прибор для регистрации движений грудной клетки, итальянский учёный А. Моссо предложил прибор для изучения кровенаполнения органов (см.Плетизмография), прибор для исследования утомления (эргограф) и весовой стол для изучения перераспределения крови. Были установлены законы действия постоянного тока на возбудимую ткань (нем. учёный Э. Пфлюгер, рус. – Б.Физиология Вериго,), определена скорость проведения возбуждения по нерву (Г. Гельмгольц). Гельмгольц же заложил основы теории зрения и слуха. Применив метод телефонического выслушивания возбуждённого нерва, рус. физиолог Н. Е. Введенский внёс значительный вклад в понимание основных физиологических свойств возбудимых тканей, установил ритмический характер нервных импульсов. Он показал, что живые ткани изменяют свои свойства как под действием раздражителей, так и в процессе самой деятельности. Сформулировав учение об оптимуме и пессимуме раздражения, Введенский впервые отметил реципрокные отношения в центральной нервной системе. Он первый начал рассматривать процесс торможения в генетической связи с процессом возбуждения, открыл фазы перехода от возбуждения к торможению. Исследования электрических явлений в организме, начатые итал. учёными Л. Гальвани и А. Вольта, были продолжены нем. учёными – Дюбуа-Реймоном, Л. Германом, а в России – Введенским. Рус. учёные И. М. Сеченов и В. Я.Данилевский впервые зарегистрировали электрические явления в центральной нервной системе. Развернулись исследования нервной регуляции физиологических функций с помощью методик перерезок и стимуляции различных нервов. Нем. учёные братья Э. Г. и Э. Вебер открыли тормозящее действие блуждающего нерва на сердце, рус. физиолог И. Физиология Цион – учащающее сердечные сокращения действие симпатического нерва, И. П. Павлов – усиливающее действие этого нерва на сердечные сокращения. А. П. Вальтер в России, а затем К. Бернар во Францииобнаружили симпатические сосудосуживающие нервы. Людвиг и Цион обнаружили центростремительные волокна, идущие от сердца и аорты, рефлекторно изменяющие работу сердца и тонус сосудов. Физиология В. Овсянниковоткрыл сосудодвигательный центр в продолговатом мозге, а Н. А. Миславский подробно изучил открытый ранее дыхательный центр продолговатого мозга. В 19 в. сложились представления о трофической роли нервной системы, т. е. о её влиянии на процессы обмена веществ и питание органов. Франц. учёный Физиология Мажанди в 1824 описал патологические изменения в тканях после перерезки нервов, Бернар наблюдал изменения углеводного обмена после укола в определённый участок продолговатого мозга («сахарный укол»), Р. Гейденгайн установил влияние симпатических нервов на состав слюны, Павлов выявил трофическое действие симпатических нервов на сердце. В 19 в. продолжалось становление и углубление рефлекторной теории нервной деятельности. Были подробно изучены спинномозговые рефлексы и проведён анализрефлекторной дуги. Шотл. учёный Ч. Белл в 1811, а также Мажанди в 1817 и нем. учёный И. Мюллер изучили распределение центробежных и центростремительных волокон в спинномозговых корешках (Белла – Мажанди закон).Белл в 1826 высказал предположение об афферентных влияниях, идущих от мышц при их сокращении в центральную нервную систему. Эти взгляды были затем развиты русскими учёными А. Фолькманом, А. М. Филомафитским. Работы Белла и Мажанди послужили толчком для развития исследований по локализации функций в мозге и составили основу для последующих представлений о деятельности физиологических систем по принципу обратной связи. В 1842 французский физиолог П. Флуранс, исследуя роль различных отделов головного мозга и отдельных нервов в произвольных движениях, сформулировал понятие о пластичности нервных центров и ведущей роли больших полушарий головного мозга в регуляции произвольных движений. Выдающееся значение для развития Физиология имели работы Сеченова, открывшего в 1862 процесс торможения в центральной нервной системе. Он показал, что раздражение мозга в определённых условиях может вызывать особый тормозной процесс, подавляющий возбуждение. Сеченовым было также открыто явление суммации возбуждения в нервных центрах. Работы Сеченова, показавшего, что «... все акты сознательной и бессознательной жизни, по способу происхождения, суть рефлексы» («Рефлексы головного мозга», см. в кн.: Избранные философские и психологические произв., 1947, с. 176), способствовали утверждению материалистической Физиология Под влиянием исследований Сеченова С. П. Боткин и Павлов ввели в Физиологияпонятие нервизма, т. е. представление о преимущественном значении нервной системы в регулировании физиологических функций и процессов в живом организме (возникло как противопоставление понятию о гуморальной регуляции). Изучение влияний нервной системы на функции организма стало традицией рус. и сов. Физиология Во 2-й половине 19 в. с широким применением метода экстирпации (удаления) было начато изучение роли различных отделов головного и спинного мозга в регуляции физиологических функций. Возможность прямого раздражения коры больших полушарий была показана нем. учёными Г. Фричем и Э. Гитцигом в 1870, а успешное удаление полушарий осуществлено Физиология Гольцем в 1891 (Германия). Широкое развитие получила экспериментально-хирургическая методика (работы В. А. Басова, Л. Тири, Л. Велла, Р. Гейденгайна, Павлова и др.) для наблюдения над функциями внутренних органов, особенно органов пищеварения, Павлов установил основные закономерности в работе главных пищеварительных желёз, механизм их нервной регуляции, изменение состава пищеварительных соков в зависимости от характера пищевых и отвергаемых веществ. Исследования Павлова, отмеченные в 1904 Нобелевской премией, позволили понять работу пищеварительного аппарата как функционально целостной системы. В 20 в. начался новый этап в развитии Физиология, характерной чертой которого был переход от узкоаналитического понимания жизненных процессов к синтетическому. Огромное влияние на развитие отечественной и мировой Физиологияоказали работы И. П. Павлова и его школы по Физиология высшей нервной деятельности. Открытие Павловым условного рефлекса позволило на объективной основе приступить к изучению психических процессов, лежащих в основе поведения животных и человека. На протяжении 35-летнего исследования высшей нервной деятельности Павловым установлены основные закономерности образования и торможения условных рефлексов, физиология анализаторов, типы нервной системы, выявлены особенности нарушения высшей нервной деятельности при экспериментальных неврозах, разработана корковая теория сна и гипноза, заложены основы учения о двух сигнальных системах. Работы Павлова составили материалистический фундамент для последующего изучения высшей нервной деятельности, они дают естественнонаучное обоснование теории отражения, созданной В. И. Лениным. Крупный вклад в исследования Физиология центральной нервной системы внёс английский физиолог Ч. Шеррингтон,который установил основные принципы интегративной деятельности мозга: реципрокное торможение, окклюзию,конвергенцию возбуждений на отдельных нейронах и т.д. Работы Шеррингтона обогатили Физиология центральной нервной системы новыми данными о взаимоотношении процессов возбуждения и торможения, о природе мышечного тонуса и его нарушении и оказали плодотворное влияние на развитие дальнейших исследований. Так, голландский учёный Р. Магнус изучил механизмы поддержания позы в пространстве и ее изменения при движениях. Сов. учёный В. М. Бехтерев показал роль подкорковых структур в формировании эмоциональных и двигательных реакций животных и человека, открыл проводящие пути спинного и головного мозга, функции зрительных бугров и т.д. Сов. учёный А. А.Ухтомский сформулировал учение о доминанте как о ведущем принципе работы головного мозга; это учение существенно дополнило представления о жёсткой детерминации рефлекторных актов и их мозговых центров. Ухтомский установил, что возбуждение мозга, вызванное доминирующей потребностью, не только подавляет менее значимые рефлекторные акты, но и приводит к тому, что они усиливают доминирующую деятельность. Значительными достижениями обогатило Физиология физическое направление исследований. Применение струнного гальванометра голландским учёным В. Эйнтховеном, а затем советским исследователем А. Физиология Самойловым дало возможность зарегистрировать биоэлектрические потенциалы сердца. С помощью электронных усилителей, позволивших в сотни тысяч раз усиливать слабые биопотенциалы, американский учёный Г. Гассер, английский – Э. Эдриан и рус. физиолог Д. С. Воронцов зарегистрировали биопотенциалы нервных стволов (см. Биоэлектрические потенциалы).Регистрация электрических проявлений деятельности головного мозга – электроэнцефалография – впервые осуществлена рус. физиологом В. В. Правдич-Неминским и продолжена и развита нем. исследователем Г. Бергером. Советский физиолог М. Н. Ливанов применил математические методы для анализа биоэлектрических потенциалов коры головного мозга. Английский физиолог А. Хилл зарегистрировал теплообразование в нерве при прохождении волны возбуждения. В 20 в. начались исследования процесса нервного возбуждения методами физической химии. Ионная теория возбуждения была предложена рус. учёным В. Ю. Чаговцем, затем развита в трудах нем. учёных Ю. Бернштейна, В. Нернста и рус. исследователя П. П. Лазарева. В работах английских учёных П. Бойла, Э. Конуэя и А. Ходжкина, А.Хаксли и Б. Каца получила глубокое развитие мембранная теория возбуждения. Советский цитофизиолог Д. Н. Насоновустановил роль клеточных белков в процессах возбуждения. С исследованиями процесса возбуждения тесно связано развитие учения о медиаторах, т. е. химических передатчиках нервного импульса в нервных окончаниях (австр. фармаколог О. Лёви, Самойлов, И. П. Разенков, А. В. Кибяков, К. М. Быков, Л. С. Штерн, Е. Б. Бабский, Х. С. Коштоянц в СССР; У. Кеннон в США; Б. Минц во Франции и др.). Развивая представления об интегративной деятельности нервной системы, австралийский физиолог Дж. Эклс подробно разработал учение о мембранных механизмах синаптической передачи. В середине 20 в. американский учёный Х. Мэгоун и итальянский – Дж. Моруцци открыли неспецифические активирующие и тормозные влияния ретикулярной формации на различные отделы мозга. В связи с этими исследованиями значительно изменились классические представления о характере распространения возбуждений по центральной нервной системе, о механизмах корково-подкорковых взаимоотношений, сна и бодрствования, наркоза, эмоций и мотиваций. Развивая эти представления, советский физиолог П. К. Анохин сформулировал понятие о специфическом характере восходящих активирующих влияний подкорковых образований на кору мозга при реакциях различного биологического качества. Детально изучены функции лимбической системы мозга (амер. учёный П. Мак-Лейн, сов. физиолог И. С. Бериташвили и др.), выявлено её участие в регуляции вегетативных процессов, в формировании эмоций и мотиваций, процессов памяти, изучаются физиологические механизмы эмоций (амер. исследователи Физиология Бард, П. Мак-Лейн, Д. Линдели, Дж. Олдс; итал. – А. Цанкетти; швейцарский – Р. Хесс, Р. Хунспергер; советский – Бериташвили, Анохин, А. В. Вальдман, Н. П. Бехтерева, П. В. Симонов и др.). Исследования механизмов сна получили значительное развитие в работах Павлова, Хесса, Моруцци, франц. исследователя Жуве, сов. исследователей Физиология П. Майорова, Н. А. Рожанского, Анохина, Н. И. Гращенкова и др. В начале 20 в. сложилось новое учение о деятельности желёз внутренней секреции – эндокринология. Были выяснены основные нарушения физиологических функций при поражениях желёз внутренней секреции. Сформулированы представления о внутренней среде организма, единой нейро-гуморальной регуляции, гомеостазе, барьерных функциях организма (работы Кеннона, сов. учёных Л. А. Орбели, Быкова, Штерн, Г. Н. Кассиля и др.). Исследованиями Орбели и его учеников (А. В. Тонких, А. Г. Гинецинского и др.) адаптационно-трофической функции симпатической нервной системы и её влияния на скелетную мускулатуру, органы чувств и центральную нервную систему, а также школой А. Д.Сперанского – влияние нервной системы на течение патологических процессов – было развито представление Павлова о трофической функции нервной системы. Быков, его ученики и последователи (В. Н. Черниговский, И. А. Булыгин, А. Д. Слоним, И. Т. Курцин, Э. Ш. Айрапетьянц, А. В. Риккль, А. В. Соловьев и др.) развили учение о кортико-висцеральной физиологии и патологии. Исследованиями Быкова показана роль условных рефлексов в регуляции функций внутренних органов. В середине 20 в. значительных успехов достигла Физиология питания. Были изучены энерготраты людей различных профессий и разработаны научно обоснованные нормы питания (сов. учёные М. Н. Шатерников, О. П. Молчанова, нем. исследователь К. Фойт, амер. физиолог Физиология Бенедикт и др.). В связи с космическими полётами и исследованиями водного пространства развиваются космическая и подводная Физиология Во 2-й половине 20 в. активно разрабатывается Физиология сенсорных систем (сов. исследователи Черниговский, А. Л. Вызов, Г. В. Гершуни, Р. А. Дуринян, швед. исследователь Р. Гранит, канад. учёный В. Амасян). Сов. исследователь А. М. Уголев открыл механизм пристеночного пищеварения. Были открыты центральные гипоталамические механизмы регуляции голода и насыщения (амер. исследователь Дж. Бробек, инд. учёный Б. Ананд и многие др.). Новую главу составило учение о витаминах, хотя необходимость этих веществ для нормальной жизнедеятельности была установлена ещё в 19 в. – работы русского учёного Н. И. Лунина. Крупные успехи достигнуты в изучении функций сердца (работы Э. Старлинга, Т. Льюиса в Великобритании; К. Уиггерса в США; А. И. Смирнова, Г. И. Косицкого, Физиология З. Меерсона в СССР; и др.), кровеносных сосудов (работы Х. Геринга в Германии; К. Гейманса в Бельгии; В. В. Парина, Черниговского в СССР; Э. Нила в Великобритании; и др.) и капиллярного кровообращения (работы дат. учёного А. Крога, сов. физиолога А. М. Чернуха и др.). Изучен механизм дыхания и транспорт газов кровью (работы Дж. Баркрофта, Дж. Холдейна в Великобритании; Д. Ван Слайка в США; Е. М. Крепса в СССР; и др.). Установлены закономерности функционирования почек (исследования англ. учёного А. Кешни, американского – А. Ричардса, и др.). Сов. физиологи обобщили закономерности эволюции функций нервной системы и физиологических механизмов поведения (Орбели, Л. И. Карамян и др.). На развитие Физиология и медицины оказали влияние работы канадского патолога Г. Селье, сформулировавшего (1936) представление о стрессе как неспецифической адаптивной реакции организма при действии внешних и внутренних раздражителей. Начиная с 60-х гг. в Физиология всё шире внедряется системный подход. Достижением сов. Физиология является разработанная Анохиным теория функциональной системы, согласно которой различные органы целого организма избирательно вовлекаются в системные организации, обеспечивающие достижение конечных, приспособительных для организма результатов. Системные механизмы деятельности мозга успешно разрабатываются рядом советских исследователей (М. Н. Ливанов, А. Б. Коган и многие др.). Современные тенденции и задачи физиологии. Одна из основных задач современной Физиология – выяснение механизмов психической деятельности животных и человека с целью разработки действенных мероприятий против нервно-психических болезней. Решению этих вопросов способствуют исследования функциональных различий правого и левого полушарий мозга, выяснение тончайших нейронных механизмов условного рефлекса, изучение функций мозга у человека посредством вживленных электродов, искусственного моделирования психопатологических синдромов у животных.
8.Вклад выдающихся отечественных ученых в становление физиологи как науки.
9. Современный этап развития нейрофизиологии как науки.
10. Общие структурные свойства нейрона, раздражимость и возбудимость.
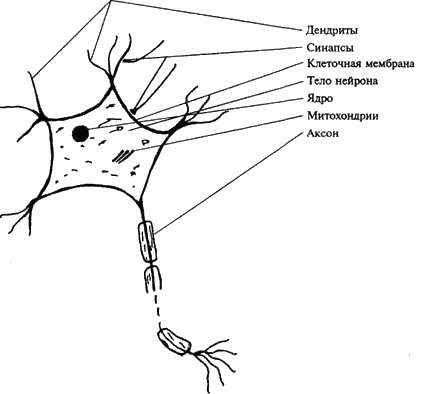
Структурно-функциональной единицей нервной системы является нервная клетка, или нейрон, или нейроцит. Всю нервную систему можно представить как взаимосвязанную и взаимодействующую сеть из нескольких триллионов нервных клеток. Несмотря на их громадное разнообразие, можно говорить о ряде общих структурных и функциональных признаков, присущих всем нервным клеткам (рис. 5). В нейроне выделяют следующие основные части: тело, отростки и их окончания. Тело нейрона, размеры которого колеблются от 4 до 130 мкм, представляет собой скопление клеточной плазмы, в которой располагается ядро – носитель генетической информации, митохондрии – универсальные «генераторы» энергии, необходимой для обеспечения деятельности клетки, и большое количество структур, выполняющих различные специфические функции. Поверхность нейрона, его оболочка, часто именуемая просто мембраной, не только обеспечивает обмен с окружающей средой, но, обладая свойствами полупроницаемой мембраны, является структурой, где развиваются сложные процессы биоэлектрогенеза, лежащие в основе главных функций нервной клетки. Отростки нервных клеток являются выростами цитоплазмы. Различают два вида отростков. Дендриты – короткие, древовидно ветвящиеся, постепенно истончаются и заканчиваются в окружающих тканях. Количество их достигает десяти, они многократно увеличивают поверхность клетки. Помимо дендритов нервная клетка всегда имеет один аксон (или нейрит). Этот отросток всегда более крупный, длинный (до 1 м) и менее ветвистый. Аксон заканчивается синапсом, при помощи которого он функционально взаимодействует с иннервируемыми структурами.
По своей функциональной значимости в составе рефлекторной дуги различают три вида нейронов:
рецепторные (чувствительные, афферентные), имеющие чувствительные нервные окончания, которые способны воспринимать раздражения из внешней или внутренней среды;
эффекторные (эфферентные), окончания аксонов которых передают нервный сигнал на рабочий орган;
ассоциативные (вставочные, центральные), являющиеся промежуточными в составе рефлекторной дуги и передающие информацию с чувствительного нейрона на эффекторные.
Следует иметь в виду, что на теле и отростках большинства нервных клеток имеется очень большое количество синапсов, через которые поступает информация с других нейронов .Несмотря на громадное морфологическое и функциональное разнообразие нейронов, можно выделить ряд ключевых свойств и функций.
К числу наиболее важных свойств относятся:
1. Наличие трансмембранной разности потенциалов, т. е. между наружной и внутренней поверхностями оболочки нейрона в покое регистрируется разность потенциала порядка 90 мВ, наружная поверхность электроположительна по отношению к внутренней. Величина и направление трансмембранного тока меняются в зависимости от состояния нейрона.
2. Очень высокая чувствительность к некоторым химическим веществам (медиаторам) и электрическому току.
3. Способность к нейросекреции, т. е. к синтезу и выделению в окружающую среду или в синаптическую щель биологически активных веществ.
4. Высокий уровень энергетических процессов, что обусловливает необходимость постоянного притока основного источника энергии – глюкозы и кислорода, необходимого для окисления.
Принято различать следующие функции нейрона:
1. Воспринимающая – эта функция представлена двумя механизмами. Во-первых, чувствительные окончания дендритов способны обеспечить рецепцию, т. е. трансформацию специфической энергии раздражителя внешней или внутренней среды в неспецифический процесс нервного возбуждения, нервный импульс, который по отростку распространяется по направлению к телу нервной клетки. Во-вторых, на всех частях нейрона имеются многочисленные (до нескольких десятков тысяч) синапсы, при помощи которых химическим путем возбуждение передается от одного нейрона к другому. Химические вещества, осуществляющие эту передачу, обозначают медиаторы (или нейротрансмиттеры). К их числу, в частности, относятся адреналин, норадреналин, дофамин, серотонин, ацетилхолин, гамма-аминомасляная кислота и многие другие. В результате воздействия медиатора в теле нервной клетки развивается возбуждение и возникновение нервного импульса или снижение возбудимости нейрона – его торможение.
2. Интегративная – обработка одновременно или в течение короткого интервала времени поступающих нервных сигналов по механизму их алгебраической суммации, в результате которой на выходе нейрона формируется сигнал, несущий в себе информацию всех суммированных сигналов.
3. Мнестическая, основанная на существовании тонких молекулярных биофизических процессов, сохраняющих след от всякого предыдущего воздействия и благодаря этому трансформирующих характер ответной реакции на всякое последующее. По существу, это элементарная форма памяти и научения.
4. Проводниковая функция, суть которой состоит в том, что от тела нейрона по аксону к его окончанию в естественных условиях только в одном этом направлении распространяется, не затухая, нервный импульс. Скорость его распространения в зависимости от морфофункциональных особенностей проводника колеблется от нескольких сантиметров до 100–120 метров в секунду.
5. Передающая, проявляющаяся в том, что нервный импульс, достигнув окончания аксона, который, собственно, уже входит в структуру синапса, обусловливает выделение медиатора – непосредственного передатчика возбуждения к другому нейрону или исполнительному органу. Часто в бытовых разговорах приходится слышать сожалеющее высказывание, что нервные клетки не восстанавливаются. Да, применительно к телу нейрона, это действительно так, и в ряде случаев это действительно плохо. Но следует также иметь в виду, что количество нейронов у человека значительно превышает его потребности на протяжении всей жизни. И, кроме того, как указывалось выше, нервные клетки на протяжении жизни человека «обучаются», «приобретают опыт», а потому включение в слаженный нейрональный ансамбль «необученного» элемента затруднило бы его работу.
Возбудимость- этой свойство нейрона генерировать ПД на раздражение. Возбудимость является частным случаем наиболее общего свойства всех клеток- раздражимости. К возбудимым клеткам относятся только те, которые генерируют ПД. Это мышечные и нервные клетки. Нередко к возбудимым тканям необоснованно относят и «железистую ткань», хотя железистой ткани нет, а имеются различные железы и железистый эпителий как вид тканей. В процессе активной деятельности железы в ней действительно регистрируется биоэлектрические явления, поскольку железа как орган состоит из различных клеток: соединительной ткани, эпителиальной, мышечной. ПД проводится по мембранам нервных и мышечных клеток, с его помощью передается информация и обеспечивается управление деятельностью клеток организма.
Невозбудимыми тканями являются эпителиальная и соединительная( собственно соединительная, ретикулярная, жировая, хрящевая, костная и т.д) клетки этих тканей не генерируют ПД при действии на них раздражителя.
Раздражи́мость- свойство живых организмов реагировать на различные воздействия (раздражители) соответствующими изменениями на уровне клетки, ткани или всего организма. Раздражимость связана с приспособлением организма к изменяющимся условиям внешней среды. У растений проявляется в реакциях на свет, гравитацию, в двигательных (как и у низших животных) реакциях (см. Тропизмы, Настии). Многоклеточным животным и человеку свойственны более точные, быстрые и разнообразные реакции в ответ на раздражение. Их обеспечивают рефлексы и высшая нервная деятельность. Часто термин «раздражимость» используют как синоним «возбудимости».
11. Механизмы возникновения электрических ответов.
12.Потенциал покоя клетки как трансмембранная разность потенциалов. Роль отечественных физиологов в установлении природы потенциала покоя клетки.
Потенциа́л поко́я (ПП) — мембранный потенциал возбудимой клетки в невозбужденном состоянии. Он представляет собой разность электрических потенциалов, имеющихся на внутренней и наружной сторонах мембраны и составляет у теплокровных от -55 до -100 мВ[1]. У нейронов и нервных волокон обычно составляет -70 мВ. Измеряется изнутри клетки. ПП формируется в два этапа.
Первый этап: создание незначительной (-10 мВ) отрицательности внутри клетки за счёт неравного асимметричного обмена Na+ на K+ в соотношении 3 : 2. В результате этого клетку покидает больше положительных зарядов с натрием, чем возвращается в неё с калием. Такая особенность работы натрий-калиевого насоса, осуществляющего взаимообмен этих ионов через мембрану с затратами энергии АТФ, обеспечивает его электрогенность.
Результаты деятельности мембранных ионных насосов-обменников на первом этапе формирования ПП таковы:
1. Дефицит ионов натрия (Na+) в клетке.
2. Избыток ионов калия (K+) в клетке.
3. Появление на мембране слабого электрического потенциала (-10 мВ).
Второй этап: создание значительной (-60 мВ) отрицательности внутри клетки за счёт утечки из неё через мембрану ионов K+. Ионы калия K+ покидают клетку и уносят с собой из неё положительные заряды, доводя отрицательность до -70 мВ.Итак, мембранный потенциал покоя - это дефицит положительных электрических зарядов внутри клетки, возникающий за счёт утечки из неё положительных ионов калия и электрогенного действия натрий-калиевого насоса.
13. Ионные каналы (транспортная система, воротный механизм)
Через отверстие в клеточной мембране способны проходить только конкретные частицы, поэтому эти отверстия называются ионными каналами. Движением потоков ионов через канал управляют ворота. Ворота бывают медленные ( не активационные) и быстрые (активационные) . активационные ворота управляются электрическим полем мембраны.
Ионные каналы (ИК) - это мембранные молекулярные структуры, образованные интегральными (трансмембранными) белками, пронизывающими клеточную мембрану поперёк в виде нескольких петель и образующими в мембране сквозной канал (пору). Канальные белки состоят из субъединиц, образующих структуру со сложной пространственной конфигурацией, в которой кроме поры обычно имеются дополнительные молекулярные системы: открытия, закрытия, избирательности, инактивации, рецепции и регуляции. ИК могут иметь не один, а несколько участков (сайтов) для связывания с управляющими веществами (лигандами).
Ионные каналы можно рассматривать как тракнспортный механизм, обеспечивающий перемещение ионов между цитоплазмной клетки и наружной средой.
Кроме ИК в мембране суцществуют и другие транспортные системы для переноса через неё различных веществ . Транспортные структуры мембраны - это специальные белковые структуры, встроенные в мембрану и обеспечивающие трансмембранный транспорт, т.е. перенос веществ через мембрану. многие вещества могут проходить сквозь клеточную мембрану самостоятельно, за счёт простой диффузии. Они "автоматически" перемещаются из зоны своей повышенной концентрации в зону пониженной концентрации, т. к. стремятся уравнять свою концентрацию по обе стороны мембраны. Так обычно ведут себя все вещества в растворах: они стремятся равномерно распределиться по всему объёму жидкости, пытаясь преодолеть разделяющую раствор на отсеки преграду. В принципе, любая молекула может пройти через липидный бислой клеточной мембраны, потому что составляющие его молекулы липидов сохраняют некоторую подвижность относительно друг друга и могут временами раздвигаться в стороны, пропуская различные вещества. Однако скорость такой пассивной диффузии, т.е. перехода вещества через мембрану из области с большей концентрацией в область с меньшей, может сильно различаться для разных веществ. Для многих веществ диффузия занимает столь длительное время, что можно говорить о практической непроницаемости для них мембраны. Скорость диффузии различных веществ через мембрану зависит главным образом от размера их молекул, электрического заряда (полярности) и их относительной растворимости в жирах. Легче и лучше всего через мембрану пассивно проникают жирорастворимые неполярные мелкие молекулы. Таким образом, свободно проникать сквозь мембрану в клетку и обратно могут толькожирорастворимые вещества, способные растворяться в жировом (липидном) слое мембраны. Транспорт других веществ через мембрану требует особых механизмов. Какие же вещества необходимо протаскивать через мембрану «насильно»? Это все полярные молекулы, не растворимые в жирах: молекулы воды, ионы (электролиты), а также более крупные молекулы питательных веществ, таких как глюкоза и аминокислоты.
Для транспорта в клетку веществ, слабо способных к диффузии через липидный слой мембраны, необходимы специальные транспортные структуры.
Виды транспортных структур мембраны:
1. Ионные каналы - это специальные поры (дырочки) в мембране, образованные канальными белками, позволяющие ионам проходить через мембрану в обоих направлениях: как внутрь, так и наружу.
2. Транслоказы, - специальные мембранные белки, облегчающие переход вещества через мембрану за счёт своего временного связывания с диффундирующим веществом. Не требуют энергии, работают в обоих направлениях в зависимости от концентрации переносимого вещества.
3. Транспортёры - белковые структуры, насильно протаскивающие определённые вещества сквозь клеточную мембрану в определённом направлении с затратами энергии. Ионные насосы - это транспортёры ионов. По способу использования энергии для своей работы транспортёры можно разделить на "симпортные" и "антипортные". Симпортные транспортёрыиспользуют совместный транспорт в одном направлении двух веществ: одно из них должно иметь большую потенциальную энергию для движения через мембрану. Например, симпорт в клетку с помощью ионов натрия глюкозы, или симпорт ионов кальция с помощью ионов натрия. Антипортные транспортёры (обменники) используют встречный транспорт двух веществ с разной потенциальной энергией диффузии. Так работает, например, натрий-калиевый ионный насос.
Итак, перенос веществ через клеточную мембрану происходит различными путями.
Механизмы транспорта веществ через мембрану
1. Простая диффузия жирорастворимых (гидрофобных) веществ через жировой слой мембраны. Это пассивный процесс под действием градиента (перепада) концентрации вещества по разные стороны мембраны. (Смотрите видео: пассивный транспорт через мембрану).
2. Неуправляемая диффузия (неуправляемый пассивный перенос) водорастворимых веществ через постоянно открытые ионные каналы мембраны.
3. Управляемая диффузия (управляемый пассивный перенос) водорастворимых веществ через управляемые ионные каналы мембраны. (Смотри: Ионные каналы мембраны)
4. Активный транспорт водорастворимых веществ с помощью специальных белковых транспортных структур (транспортёров) за счёт использования энергии расщепления АТФ. Одна из самых главных транспортных структур мембраны — это фермент АТФаза
14. Пассивный ионный транспорт.
Существуют два основных вида транспорта ионов и молекул через мембрану: активный и пассивный транспорт.Пассивный транспорт — это движение частиц через мембрану в направлении концентрационного градиента, а для заряженных частиц (в частности, ионов) — еще и электрического градиента. Такой транспорт не требует затрат энергии.
Структуры, обеспечивающие транспорт ионов
Пассивный транспорт
Липидный бислой, составляющий основу клеточных мембран, не проницаем для заряженных частиц. Поэтому ионы могут диффундировать сквозь мембрану только через специальные трансмембранные (пронизывающие мембрану) белковые комплексы —ионные каналы. Эти комплексы образуют стенки поры, сквозь которую и проходят ионы. Напоминаем, что в данном случае речь идет о простой диффузии. Пассивный транспорт Диффузия может происходить либо через липидный бислой, или при участии белков. Диффузия через липидный бислой - это простая диффузия, зависит от концентрации веществ по обе стороны мембраны и температуры. Так переходят через мембрану вещества, которые растворяются в липидах (О2, СО2, азот, алкоголь). Кислород проходит так легко, будто мембраны и не существует. Через мембрану легко проходит и вода благодаря небольшому размеру ее молекул. Простая диффузия происходит не только через липидный бислой, но и через белковые каналы и «ворота» в них. Эти каналы высокоселективные, транспорт через них зависит от вида ионов или молекул, их диаметра, формы, электрического заряда. Натриевые каналы имеют диаметр 0,3-0,5 нм, их внутренняя поверхность отрицательно заряжена. К +-канал имеет диаметр 0,3 нм, электрического заряда не имеет. Конформационные изменения формы белковых молекул влияют на состояние «ворот» (открытые или закрытые), в Na +-каналам они расположены на внешней, а в К +-каналах - на внутренней поверхности канала. Состояние «ворот» контролируется двумя принципиально отличными механизмами: 1) состояние «ворот» зависит от разности электрических потенциалов по обе стороны мембраны, это потенциалзависимые каналы, 2) хемочутливи каналы те, белковые соединения которых способны связываться с некоторыми веществами-лигандами. Это вызывает конформационные изменения белковой молекулы и реакцию «ворот». К лиганд принадлежит ацетилхолин. Потенциалчутливи каналы реагируют по закону, формулирует ться словосочетанием «все или ничего». Состояние «ворот» меняется очень быстро (миллионные доли секунды). Сейчас есть точные методики исследования потока ионов через одиночные каналы (с помощью микропипеток, диаметр кончика которых составляет 1-2 мкм). Облегченная диффузия происходит при участии специфического белка-переносчика, что способствует диффузии. От простой диффузии отличается тем, что скорость ее увеличивается только до определенной степени, а затем остается постоянной. Таким образом транспортируются глюкоза и большинство аминокислот. На скорость диффузии влияют концентрация вещества по обе стороны мембраны,-ее проницаемость и ряд других факторов. А проницаемость зависит от толщины мембраны, количества каналов на единицу площади, молекулярной массы вещества, температуры. Осмос имеет значение для транспорта воды через мембраны, проницаемые для воды, но не для ионов (полупроницаемые мембраны). Если концентрация вещества по обе стороны мембраны различна, то она будет переходить туда, где концентрация выше. Это наблюдается, пока концентрация не выровняется. Для создания осмотического давления имеет значение не молекулярная масса вещества, а количество молекул на единицу объема раствора. Переход воды по законам осмоса происходит очень быстро. Определяется величина осмотического давления в атмосферах, в миллиметрах ртутного столба или в милиосмолях. Эта величина в клеточной и внеклеточной жидкости составляет 7,6 атм, или 300 мосм / кг