

§ 26. Класс Костные рыбы
Общая характеристика класса Костные рыбы. Внутри типа хордовых это самый многочисленный класс, насчитывающий более 22 тыс. видов. Костные рыбы населяют все водоемы земного шара. Разнообразие условий жизни в разных водоемах обусловило морфофизиологическое и экологическое многообразие представителей этого класса. Однако, несмотря на широкую дивергенцию, все виды костных рыб обладают общими признаками и особенностями организации.
Тело рыб покрыто костной чешуей, которая имеет разные типы строения, но никогда не бывает плакоидной. Скелет костный, у примитивных костных рыб иногда сохраняется хрящ, но при этом всегда имеются покровные костные образования. В черепе формируются новообразования — вторичные челюсти с зубами и жаберная крышка, закрывающая снаружи жаберный аппарат. Межжаберные перегородки редуцированы, жаберные лепестки располагаются непосредственно на жаберных дугах. У большинства видов образуется плавательный пузырь как вырост на спинной стороне пищевода. В сердце отсутствует артериальный конус, у большинства имеется расширение передней части брюшной аорты — луковица аорты. Оплодотворение в основном наружное. Икра не имеет плотных
оболочек. Обычно самка откладывает очень большое количество икры (например, рыба-луна — до 300 млн икринок). В связи с разными условиями обитания форма тела, подвижность и пищевая специализация костных рыб варьируют в очень широких пределах.
Рыбы, живущие в толще воды, как правило, имеют сплющенную с боков веретеновидную форму. У активных пловцов (скумбрий, тунцов) она торпедообразная. У стремительных спринтеров тело стреловидное (щука, сайра). Встречаются рыбы с шаровидным (иглобрюх), змеевидным (угорь), лентовидным (мурена) (рис. 81), дисковидным (рыба-луна) телом. У донных рыб тело сплющено в спинно-брюшном направлении.
Костных рыб разделяют на 4 подкласса: Хрящекостные, Кистеперые, Двоякодышащие, Лучеперые2.
Подкласс Хрящекостные, или Хрящевые ганоиды.
К этой группе относятся древние рыбы, обладающие многими примитивными чертами строения. По внешнему виду они напоминают хрящевых рыб: хвост гетероцеркальный, имеется рострум, рот в виде щели находится на нижней поверхности головы. Парные плавники располагаются горизонтально по отношению к телу. Тело покрыто костной чешуей, некоторые чешуи срастаются, образуя костные пластинки — жучки. Хорда сохраняется в течение всей жизни. Тела позвонков не формируются, но развиваются хрящевые верхние и нижние дуги. Череп практически целиком хрящевой, но поверх него развивается костный панцирь и жаберная крышка. Как и у акул, в кишечнике находится спиральный клапан. В сердце развит артериальный конус. Плавательный пузырь сохраняет связь с кишечником. Оплодотворение наружное, икра мелкая. Большинство видов — проходные: живут в морях, а на размножение поднимаются в реки. Питаются животной пишей (рыбами, моллюсками, насекомыми). Представители: осетровые (осетр, белуга, стерлядь, севрюга и др.) и веслоносые. Имеют важное промысловое значение.
Подкласс Двоякодышащие рыбы. Немногочисленная группа пресноводных рыб, совмещающая примитивные признаки предковых форм с прогрессивными приспособлениями к обитанию в обедненных кислородом водоемах. Тела позвонков у них не развиваются, осевым скелетом на протяжении всей жизни является хорда. Основания плавников имеют вид мясистых лопастей, покрытых чешуей. С их помощью рыбы могут не только плавать, но и передвигаться по дну.
Двоякодышащие рыбы обитают в пресных, бедных кислородом тропических водоемах. Дыхание жаберное и легочное. С брюшной стороны пищевода имеются 1—2 полых выроста, выполняющих роль легких. В сердце намечается разделение предсердия и формирование второго круга кровообращения. При недостатке кислорода в воде или во время спячки дыхание только легочное. Современные представители: однолегочные — австралийский рогозуб и двулегочные — чешуйчатники (африканские протоптеры и южноамериканский лепидосирен). Рогозубы живут в непересыхающих водоемах и в спячку не впадают. Чешуйчатники при пересыхании водоемов могут зарываться в грунт и на долгий период (до 9 месяцев) впадать в спячку. Протоптер при этом даже образует капсулу.
Подкласс Кистеперые рыбы. Кистеперые рыбы долгое время считались вымершей группой. В 1938 г. был обнаружен единственный современный вид — латимерия, который обитает в районе Коморских островов на глубине около тысячи метров. Кистеперые близки к двоякодышащим и произошли, видимо, от общего предка. Особенность кистеперых рыб — наличие мускулатуры в составе конечностей и расчлененность их скелета. В эволюции это стало предпосылкой для превращения плавников в пятипалые конечности. Древние кистеперые рыбы обитали в пресных водоемах и имели двойное дыхание: при недостатке кислорода они поднимались на поверхность и дышали воздухом. Их развитие шло в двух направлениях: одна ветвь дала начало предкам современных земноводных, а другая приспособилась к жизни в морской воде. Современная латимерия, в отличие от своих мезозойских предков, не способна к дыханию атмосферным кислородом, ее большое дегенерировавшее легкое заполнено жиром.
Подкласс Лучеперые рыбы. Этот подкласс объединяет большинство ныне живущих рыб. Тело покрывает костная чешуя — тонкие пластинки, черепицеобразно налегающие друг на друга. Скелет костный, иногда сохраняется небольшое количество хрящевой ткани. Хвостовой плавник обычно гомоцеркальный (равнолопастной). Парные плавники расположены вертикально по отношению к телу, а не горизонтально, как у хрящевых рыб. Лопасти плавников поддерживаются костными лучами, за что подкласс и получил свое название — Лучеперые. У большинства видов в сердце отсутствует артериальный конус. Кишечник удлиняется, а спиральный клапан редуцируется. Клоака отсутствует, кишечник заканчивается анальным отверстием. Этот подкласс включает около 40 современных отрядов (Сельдеобразные, Окунеобразные, Карпообразные, Лососеобразные и др.).
Рассмотрим организацию костных рыб на примере представителя надотряда Костистые рыбы — речного окня, типичного обитателя пресноводных водоемов.
Внешнее строение и покровы. Тело окуня имеет обтекаемую форму, покрыто костной чешуей. Эти чешуи образуются в соединительнотканном слое кожи и растут на протяжении всей жизни рыбы: в течение года на чешуйке образуется новое кольцо. Снаружи чешуя покрыта многослойным эпителием, в котором располагаются многочисленные одноклеточные железы. Эти железы вырабатывают слизь, которая уменьшает трение при передвижении в воде. Тело рыбы делится на голову, туловище и хвост, заканчивающийся хвостовым плавником. Благодаря движению хвостового плавника и изгибанию туловища рыба движется вперед. Кроме хвостового, у рыб имеются парные брюшные и грудные плавники, а также непарные плавники —- один или несколько спинных и подхвостовой (анальный). При помощи парных плавников поддерживается нормальное положение рыбы спиной вверх, а также осуществляется подъем и спуск на нужную глубину. Спинной и анальный плавники придают рыбе устойчивость.
Опорно-двигательная система. Скелет костной рыбы разделяется на осевой скелет, череп (мозговой и висцеральный), скелет парных плавников и их поясов и скелет непарных плавников (рис. 82).
Осевой скелет представлен позвоночником, образованным двояковогнутыми позвонками. Сильно редуцированная хорда имеет четкообразное строение: она расширяется в пространстве между телами позвонков и сильно сужается при прохождении через канал в центре тела позвонка.

Позвоночник делится на два отдела: туловищный и хвостовой. Костные позвонки имеют верхние и нижние дуги. Верхние дуги позвонка, срастаясь, образуют позвоночное отверстие. Отверстия всех позвонков формируют позвоночный канал, в котором проходит спинной мозг. После срастания верхние дуги продолжаются в непарный верхний остистый отросток. Снизу от позвонков отходят нижние дуги. В туловищном отделе к ним прикрепляются ребра. В отличие от хрящевых рыб, ребра которых ограничивают полость тела лишь сверху, у костных ребра длиннее и ограничивают полость тела еще и с боков. Нижние дуги хвостовых позвонков смыкаются, образуя гемальный канал, в котором проходят хвостовые артерия и вена.
Для каждого вида рыб характерно определенное общее число позвонков, например у угря их 114, у северной сельди — 57, а у рыбы-луны — всего 17. Интересно, что повышение температуры окружающей среды в период эмбрионального развития приводит к уменьшению числа позвонков.
Череп почти полностью костный. Кости мозгового черепа соединяются между собой неподвижно и защищают головной мозг и органы чувств. Через затылочное отверстие в мозговом черепе происходит соединение головного и спинного мозга. Висцеральный череп состоит из висцеральных дуг (челюстной, подъязычной и жаберных) и жаберной крышки.
Пояс передних конечностей (плечевой пояс) соединен с мозговым черепом. Скелет грудных плавников (передних конечностей) присоединен к поясу передних конечностей. Пояс задних конечностей (тазовый пояс) лежит в толще мускулатуры и не имеет опоры на позвоночник. К нему причленяется скелет брюшных плавников (задних конечностей). Скелет парных и непарных плавников образован костями и многочисленными костными лучами, поддерживающими лопасть плавников.
Мускулатура рыб сохраняет правильную сегментацию на спинной стороне тела, где расположены мышечные сегменты (миомеры), разделенные перегородками (миосептами). Часть сегментов дифференцировались и образовали отдельные группы мышц: глазные, челюстные, жаберные, мышцы парных плавников.
Пищеварительная система. В ротовой полости большинства рыб на челюстях располагаются конические зубы. I) течение всей жизни происходит нерегулярная смена зубов: новые зубы вырастают между старыми. Строение челюстей и зубов зависит от типа и способа питания. Из роговой полости пища попадает в глотку. За глоткой начинается мускулистый пищевод, который переходит в желудок (рис. 83). У некоторых рыб (карповые, многие бычки) желудок отсутствует. В начальной части кишечника располагаются пилорические придатки, их число варьирует. Пилорические придатки увеличивают поверхность кишечника, в них вырабатываются пищеварительные ферменты и начинается всасывание переваренной пищи. Кишечник относительно длинный, образует петли. В передний отдел тонкого кишечника (двенадцатиперстную кишку) впадает желчный проток печени. Поджелудочная железа у большинства костистых рыб в виде небольших долек разбросана на брыжейке средней кишки. Кишечник заканчивается анальным отверстием.
У большинства костных рыб имеется плавательный пузырь. У лучеперых рыб он образуется как вырост спинной стороны пищевода. Первоначальное заполнение плавательного пузыря газом происходит за счет заглатывания мальком атмосферного воздуха. Плавательный пузырь выполняет гидростатическую функцию — изменение объема газов в пузыре приводит к изменению плотности тела рыбы. Плавательный пузырь также участвует в газообмене, служит органом, воспринимающим изменения давления, а у некоторых рыб участвует в произведении звуков.

У закрытопузырных рыб во взрослом возрасте связь пузыря с пищеводом утрачивается, а у открытопузырных — сохраняется в течение всей жизни. У закрытопузырных рыб изменение объема плавательного пузыря происходит в результате газообмена в сети капилляров, оплетающих пузырь, у открытопузырных — за счет его сжатия и расширения.
Дыхательная система. Органы дыхания — жабры эктодермального происхождения. Межжаберные перегородки отсутствуют, жаберные лепестки располагаются прямо на жаберных дугах, их свободные концы свисают в жаберную полость, ограниченную снаружи жаберной крышкой. При вдохе вода через ротовое отверстие входит в глотку, проходит через жаберные щели и попадает в жаберную полость, где омывает жаберные лепестки. Газообмен между водой и кровью происходит в многочисленных капиллярах жаберных лепестков. При выдохе вода выходит наружу из-под жаберных крышек. На внутренней стороне жаберных дуг находятся жаберные тычинки. Они образуют цедильный аппарат, который препятствует выбросу пищи наружу из глотки через жаберную полость. Наличие жаберной крышки повышает эффективность дыхательных движений.
Дыхательный акт костных рыб состоит из двух этапов. На первом из них рыба приподнимает жаберные крышки, что создает в околожаберном пространстве пониженное давление. В результате свежая вода поступает через ротовое отверстие. На втором этапе отодвинутые жаберные крышки с силой прижимаются к телу, и возросшее давление в околожаберной полости выталкивает оттуда воду во внешнюю среду. При этом вода не выходит изо рта, поскольку ротовое отверстие в тот момент закрывается.
Кровеносная система. У лучеперых костистых рыб двухкамерное сердце (одно предсердие и один желудочек) и один круг кровообращения (рис. 84). К предсердию примыкает объемная венозная пазуха, в которую собирается кровь из вен. Кровь в сердце рыб только венозная. От желудочка отходит мощная брюшная аорта. Она делится на 4 пары жаберных артерий, которые подходят к жаберным лепесткам, где ветвятся на капилляры. После газообмена обогащенная кислородом кровь впадает в парные корни спинной аорты, расположенные на спинной стороне тела. Они посылают вперед сонные артерии, снабжающие кровью головной мозг, а сзади сливаются в непарную спинную аорту. От спинной аорты отходят многочисленные сосуды, несущие богатую кислородом кровь ко всем органам тела. В хвостовом отделе спинная аорта переходит в хвостовую артерию. Венозная кровь из хвостового отдела собирается по хвостовой вене. Подходя к почкам, вена раздваивается и входит в почки, образуя воротную систему только в левой почке. Выходящие из почек задние кардинальные вены несут кровь вперед. От переднего конца тела кровь собирается по передним кардинальным венам. Передние и задние
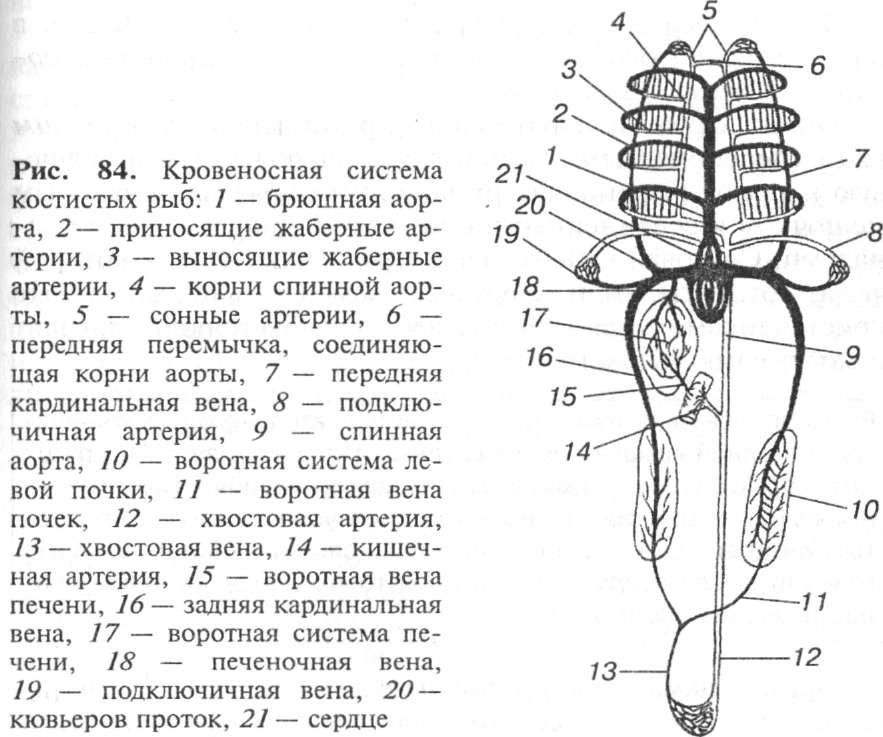
кардинальные вены сливаются в парные протоки, впадающие в венозную пазуху. Кровь от кишечника собирается в воротную вену печени, проходит через воротную систему печени и по печеночной вене тоже впадает в венозную пазуху.
Нервная система и органы чувств. Головной мозг костных рыб во многом примитивнее, чем у хрящевых рыб. Головной мозг окуня невелик по объему, он состоит из пяти отделов: переднего, промежуточного, среднего, продолговатого мозга и мозжечка. Передний мозг маленький, кора отсутствует. Наибольшего развития достигает средний мозг, сверху он разделен на две зрительные доли и полностью закрывает промежуточный мозг. У подвижных рыб хорошо развит мозжечок, у малоподвижных его относительные размеры уменьшаются. От головного мозга отходит 10 пар черепных нервов. Продолговатый мозг переходит в спинной мозг, от которого отходят спинномозговые нервы.
У рыб хорошее обоняние. Органы обоняния — обонятельные мешки с хорошо развитыми складками обонятельного эпителия.
Зрение рыб — важное средство ориентации. Видеть в воде позволяет особое строение глаза — уплощенная роговица и круглый хрусталик.
Орган слуха и равновесия представлен внутренним ухом (перепончатым лабиринтом), заключенным в хрящевую капсулу. Перепончатый лабиринт образован овальным мешочком и соединенными с ним тремя полукружными каналами. Слуховая капсула заполнена жидкостью, в которой лежат отолиты, или слуховые камешки, образованные кристалликами углекислой извести. Рыбы хорошо слышат и могут сами издавать звуки.
Рыбы производят звуки при помощи плавательного пузыря, костей жаберной крышки, лучей грудных плавников, зубов и других структур. Особенно важна звуковая коммуникация у глубоководных рыб, которые постоянно находятся в условиях полной темноты. Интересно, что рыбы способны слышать звуки, которые генерируются в воздухе, несмотря на то, что в воду при этом попадает всего 0, 01% звуковых волн.
Органы вкуса — вкусовые почки, расположенные в ротовой полости и рассеянные по всей поверхности тела.
Органы осязания — осязательные тельца в коже, особенно многочис- лены они на губах, усиках и лучах плавников.
Органы боковой линии имеют типичное строение — это погруженные в кожу каналы, на стенках которых располагаются клетки, воспринимающие давление и направление тока воды (сейсмосенсорная система).
Выделительная система. Выделительная система, как и у хрящевых рыб, представлена длинными лентовидными туловищными почками, расположенными по бокам позвоночника над плавательным пузырем. От почек отходят мочеточники. Мочеточники соединяются в выводной канал, который открывается на конце мочеполового сосочка (рис. 85). У некоторых видов имеется мочевой пузырь.
Половая система, размножение и развитие. Половая система костных рыб абсолютно не похожа на половую систему хрящевых рыб. Парные половые железы расположены по бокам от плавательного пузыря. Яичники самок имеют зернистую структуру, их задние отделы выполняют функцию выводных протоков. Таким образом, у самок костных рыб яйцеводы (мюллеровы каналы) редуцированы. Созревшие яйца (икринки) выходят либо в мочеполовой синус, либо прямо наружу через самостоятельное непарное половое отверстие. У самцов гладкие белые семенники, задние отделы которых преобразованы в семяпроводы. В отличие от хрящевых, семявыносящие протоки костных рыб самостоятельные и не имеют никакого отношения к вольфовым протокам, которые у них служат только мочеточниками. Половое отверстие у самцов и самок оканчивается на мочеполовом сосочке. Большинство видов рыб раздельнополые, однако для некоторых видов известен гермафродитизм (каменные окуни, морские караси).

Икра мелкая, покрыта студенистой оболочкой. Оплодотворение наружное. Развитие обычно идет с прохождением личиночной стадии. Из оплодотворенной икринки развивается личинка, которая питается запасами пищи, содержащимися в желточном мешке. Рот у личинки не прорывается. Спустя некоторое время личинка превращается в малька, который уже питается самостоятельно.
Плодовитость рыб гораздо выше плодовитости наземных позвоночных. Это связано с высокой смертностью — значительная часть икринок и мальков поедается хищниками. У некоторых видов рыб существует забота о потомстве, что снижает гибель икры и молодняка. Плодовитость таких рыб сокращается (трехиглая колюшка, морской конек, цихлиды и др.).
Значение рыб. В водных биогеоценозах рыбы являются обязательным звеном многих цепей питания, поэтому их роль в круговороте веществ и энергии исключительно велика. Многие виды рыб питаются донными беспозвоночными, зоо- и фитопланктоном. Крупные рыбы поедают более мелких рыб. В свою очередь, рыбы и их мальки служат пищей для головоногих моллюсков, крупных ракообразных, иглокожих, паразитических червей и других беспозвоночных. Также рыбу и ее икру используют в пищу представители всех классов позвоночных животных.
Большое значение имеют рыбы и в жизни человека. В разных странах на долю рыбы в пищевом рационе приходится от 17 до 85%. Из рыбы получают витамины (например, рыбий жир — известный витаминный препарат, богатый витамином А), кормовую муку (подкормка скоту), удобрения. Основную долю рыбы (около 90%) добывают в морях. Применяется целый ряд мер, способствующих охране и воспроизведению рыб. Действуют межправительственные соглашения о способах, объемах и сроках лова, развивается прудовое хозяйство, ведется работа по акклиматизации рыб в новых местах обитания. Декоративные и аквариумные виды рыб имеют эстетическое значение. Широкое развитие получило спортивное рыболовство.
§ 27—28. Класс Земноводные, или Амфибии. Общая характеристика и многообразие
Общая характеристика класса Земноводные. Класс Земноводные, или Амфибии, — первый класс наземных позвоночных, не утративший еще тесной связи с водной средой. Эта группа занимает промежуточное положение между водными и настоящими наземными позвоночными, что отражено в ее названии. Большинство амфибий на разных стадиях жизненного цикла обитает то в воде, то на суше, поэтому в их строении присутствуют черты и водных, и наземных позвоночных.
Большинство ученых предками амфибий считают ихтиостегий, произошедших в каменноугольном периоде от девонских кистеперых рыб. Первые настоящие земноводные, стегоцефалы, имели костный панцирь, покрывавший всю черепную коробку. Они вытеснили пресноводных кистеперых рыб из мелководных водоемов. До появления пресмыкающихся (в середине каменноугольного периода) стегоцефалы были господствующей группой в наземной фауне.
Перечислим основные прогрессивные признаки, которые позволили предкам современных амфибий освоить наземную среду:
формирование пятипалой конечности рычажного типа, которая позволяет передвигаться по твердому субстрату;
потеря жаберного дыхания и образование нового органа воздушного дыхания — легких, расположенных в полости тела;
в связи с появлением легочного дыхания — возникновение второго круга кровообращения;
прогрессивное развитие нервной системы и совершенствование органов чувств, что позволило эффективно приспособиться к новым наземным условиям существования.
Однако в строении и организации амфибий сохраняется целый ряд примитивных черт:
оплодотворение и развитие икры происходят в водной среде; вылупившаяся личинка ведет водный образ жизни и лишь в результате превращения (метаморфоза) приобретает признаки, характерные для наземных позвоночных;
взрослые амфибии имеют признаки водных животных (проницаемая для воды и газов голая кожа, несущая большое количество слизистых желез; органы выделения — туловищные почки, которые служат для выведения избытка воды из организма);
холоднокровность — температура тела амфибий зависит от температуры окружающей среды.
Морфофизиологический обзор. Рассмотрим основные характерные морфофизиологические особенности этого класса на примере травяной лягушки.
Внешнее строение и образ жизни. Травяная лягушка имеет короткое и широкое туловище, которое постепенно переходит в плоскую голову. Шея не выражена, хвоста нет. Над большим ртом расположены ноздри, а чуть выше их — крупные выпуклые глаза с веками. Ноздри снабжены особыми клапанами, закрывающими вход в легкие при погружении в воду. Позади глаз находятся крупные округлые пятна тонкой кожи — барабанные перепонки, прикрывающие снаружи полости среднего уха. Лягушка опирается на 2 пары конечностей, задние — очень длинные, между их пальцами имеется плавательная перепонка. Животное хорошо плавает, а по суше передвигается прыжками. Тонкая гладкая кожа не может защитить от высыхания, поэтому травяная лягушка обитает во влажных местах. Активна она только во влажную и теплую погоду, выходит на охоту в сумерках или после дождя. Бесхвостые амфибии, имеющие более тесную связь с водой (озерная и прудовая лягушки, жерлянки), активны в течение всего светлого времени суток.
Покровы. Кожа у лягушек голая, лишенная роговых образований. Как и у всех позвоночных, она состоит из многослойного эпидермиса и соединительнотканного кориума, или собственно кожи. В кориуме находится множество многоклеточных желез (вспомните, что у рыб одноклеточные железы располагаются в эпидермисе). Выделяемая ими слизь увлажняет кожу, предохраняя ее от высыхания. Секрет кожных желез может содержать раздражающие или ядовитые вещества (жерлянки, жабы, саламандры), защищающие животных от хищников. Проникающая через покровы вода накапливается в лимфатических полостях, расположенных под кожей и служащих резервуаром влаги при жизни на суше.
В собственно коже проходит большое количество мелких кровеносных сосудов, через стенки которых осуществляется газообмен при кожном дыхании.
Опорно-двигательная система. Выходя на сушу, животные должны были освоить новый тип передвижения по субстрату и научиться преодолевать действие сил гравитации. В результате этого скелет и мышечная система амфибий существенно изменились.
Скелет лягушки подразделяется на осевой скелет (позвоночник), скелет черепа, свободных конечностей и их поясов (рис. 86). В осевом скелете позвонки практически полностью вытесняют хорду. Позвоночник делится на четыре отдела. По сравнению с рыбами появляются два новых отдела: шейный и крестцовый. Шейный отдел состоит из одного позвонка, который обеспечивает подвижность головы в вертикальной плоскости. Число позвонков в туловищном отделе варьирует, у лягушки их семь. Единственный крестцовый позвонок несет длинные поперечные отростки, к которым причленяются кости тазового пояса. Позвонки хвостового отдела у лягушки слиты в единую кость — уростиль. У лягушки ребра не развиваются.
Выход позвоночных на сушу привел к появлению пятипалой конечности наземного типа. Скелет свободной конечности амфибий типичен для всех наземных позвоночных и при этом существенно отличается от скелета конечностей костных рыб. Конечности наземных позвоночных представляют собой рычаги, соединенные суставами. Свободная конечность состоит из трех отделов: I — плечо или бедро, II — предплечье или голень, III — кисть или стопа (соответственно для передней или задней конечности). Скелет первого отдела конечности представлен одной костью. В передней конечности — это плечевая, в задней — бедренная кость. Предплечье и голень содержат по две кости. В передней конечности — лучевая и локтевая, в задней — большая и малая берцовые. Кисть состоит из запястья, пясти и фаланг четырех-пяти пальцев. Запястье содержит около 10 мелких косточек, расположенных в три ряда, пясть — 5 удлиненных костей. Стопа состоит из предплюсны (около 10 косточек), плюсны (5 удлиненных костей) и 5 пальцев с разным числом фаланг.
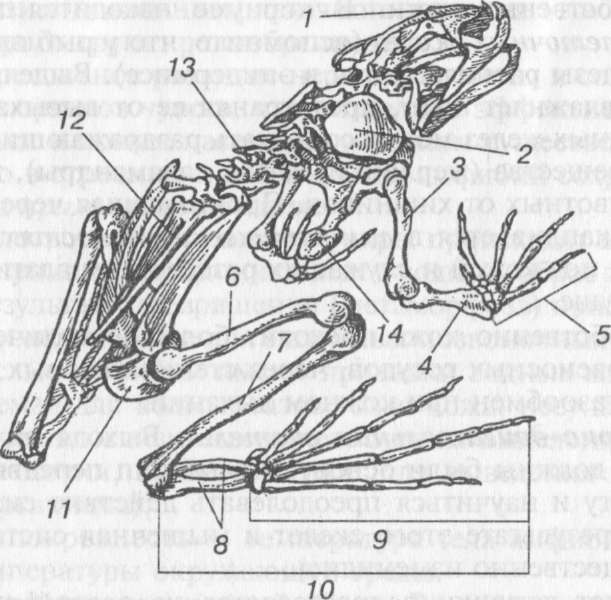
Рис. 86. Скелет лягушки: / — череп, 2 — лопатка, 3 — плечо, 4 — предплечье, 5 — кисть, 6 — бедро, 7 — голень, 8 — предплюсна, 9 — плюсна и фаланги пальцев, 10 — стопа, 11 — тазовый пояс, 12 — уростиль, 13 — позвоночник, 14 — грудина
В связи со специализацией конечностей у разных групп наземных позвоночных сильно меняется количество косточек, особенно в кисти и стопе. У лягушки в связи с приспособлением к передвижению прыжками задние конечности сильно удлиняются. Длинной становится бедренная кость, большая и малая берцовые кости срастаются в одну кость голени. Две верхние косточки предплюсны сильно увеличиваются в размерах и удлиняются, образуя так называемую пряжку, остальные остаются мелкими или редуцируются. Пять косточек плюсны, так же как и фаланги пяти пальцев, сильно удлинены. В передней конечности сливаются локтевая и лучевая кости. В кисти редуцируется первый палец, в стопе появляется рудимент добавочного пальца.
Пояс передних конечностей (плечевой пояс) имеет форму дуги, обращенной вершиной к брюшной поверхности животного. Каждая половина дуги (левая и правая) состоит из лопатки и коракоида (вороньей кости), которые образуют суставную впадину для головки плеча. Впереди от места соединения правого и левого коракоидов находится пред- грудина, а сзади — грудина. У бесхвостых между предгрудиной и лопаткой, справа и слева, расположены тонкие палочковидные ключицы. В отличие от костных рыб, плече- вой пояс амфибий лежит свободно в толще мускулатуры и не связан с черепом.
Тазовый пояс образован тремя парными элементами, которые, соединяясь между собой, образуют вертлужную впадину — место присоединения бедренной кости. У лягушек тазовый пояс состоит из длинных подвздошных костей, которые причленяются к крестцовому отделу позвоночника, сросшихся седалищных костей и лобковых хрящей.
Череп амфибий широкий и плоский, в нем сохраняется много хряща. Крупные отверстия глазниц располагаются сверху, а не с боков. Черепная коробка относительно мала. Верхняя челюсть прирастает к черепной коробке. Подъязычная и жаберные дуги, характерные для рыб, преобразуются частично в косточку среднего уха (стремечко), частично в подъязычный аппарат. Череп подвижно присоединяется к позвоночнику с помощью двух затылочных мыщелков — суставных поверхностей, расположенных по бокам от большого затылочного отверстия.
Мускулатура земноводных существенно отличается от мышечной системы рыб. Метамерность, характерная для мускулатуры рыб, выражена слабо (в туловищной мускулатуре). Большая часть мускулатуры сложно дифференцируется. У лягушек значительного развития достигает мускулатура передних и задних конечностей, что связано с передвижением по твердому субстрату. Усложняется и специализируется мускулатура ротовой полости, которая участвует в захвате и проглатывании добычи и обеспечивает дыхательные движения.
Питание и пищеварительная система. Все современные земноводные во взрослом состоянии плотоядны Они питаются различными мелкими беспозвоночными: насекомыми и их личинками, ракообразными, червями, моллюсками и другими животными. Водные виды питаются мальками рыб, самые крупные ловят других земноводных и даже птенцов водоплавающих птиц.
Ротовое отверстие ведет в обширную ротоглоточную полость, переходящую в пищевод. В ротоглоточную полость открываются парные хоаны (внутренние ноздри), евстахиевы трубы, ведущие в полость среднего уха, и гортанная щель. Мелкие одинаковые конические зубы располагаются на челюстях, они помогают удерживать добычу. У бесхвостых амфибий зубы частично редуцируются, а у жаб полностью отсутствуют. У амфибий есть слюнные железы, которые вырабатывают слизистый секрет. Этот секрет служит для смачивания пищи и не содержит пищеварительных ферментов. На дне ротовой полости находится мускулистый язык. Его форма и способы прикрепления разнообразны и зависят от типа питания. У лягушки язык прикреплен ко дну рта, а его свободная часть в спокойном состоянии обращена внутрь. При ловле добычи он выбрасывается изо рта. У всех амфибий железы слизистой оболочки выделяют липкий секрет, благодаря которому добыча приклеивается к языку. Смоченная слюной пища подается в пищевод сокращением мышц ротоглоточной полости. В акте глотания принимают участие глазные яблоки: с помощью специальных мышц они втягиваются в ротоглоточную полость и помогают проталкивать пищевой комок в пищевод.
Короткий пищевод ведет в желудок, где пища обрабатывается ферментами желудочного сока (рис. 87). От желудка отходит двенадцатиперстная кишка, в нее впадают протоки желчного пузыря и поджелудочной железы. В тонком кишечнике происходит окончательное переваривание пищи и всасывание питательных веществ. Длинный тонкий кишечник образует несколько петель и переходит в толстую кишку, которая заканчивается прямой кишкой. В толстой кишке всасывается большая часть воды и происходит формирование каловых масс. Прямая кишка открывается в клоаку.
Все отделы кишечника, а также печень и поджелудочная железа прикрепляются к спинной стороне полости тела с помощью брыжейки. В брыжейке тонкого кишечника располагается орган кроветворения — селезенка.
Дыхание и дыхательная система. Дыхательная система амфибий обеспечивает газообмен в воздушной среде и поэтому принципиально отличается от дыхательной системы рыб. Характерной особенностью земноводных является множественность органов дыхания. В поглощении кислорода и выделении углекислого газа участвуют легкие, кожа и слизистая оболочка ротоглоточной полости. Легкие представляют собой тонкостенные мешки, оплетенные сетью кровеносных сосудов. Трахея отсутствует, каждое легкое самостоятельно крепится к гортанно-трахейной камере. Наружные ноздри амфибий снабжены клапанами и открываются в ротовую полость хоанами. Газообмен в легких осуществляется за счет движения дна ротоглоточной полости. При его опускании воздух через открытые ноздри засасывается в ротоглоточную полость. Клапаны в ноздрях закрываются, открывается гортанная щель, и воздух из легких выталкивается в ротовую полость, где смешивается с находящимся там атмосферным воздухом. Затем дно полости постепенно поднимается, и смешанный воздух через гортанную шель нагнетается в легкие. После этого гортанная шель закрывается, и остатки воздуха через ноздри выталкиваются наружу. Легочное дыхание на 35—75% обеспечивает поступление в организм кислорода.

Рис. 235. Внутреннее строение лягушки:
1 — сердце; 2 —легкое; 3, 4 — печень; 5 — желчный пузырь; 6 -- желудок; 7 — поджелудочная железа; 8, 9 — топкая кишка; 10 — толстая кишка; 11 -- селезенка; 12 — клоака; 13 — мочевой пузырь; 14—его отверстие в клоаке; 15— почка; 16 - мочеточник;
17 — его отверстие в клоаке; 18 — яичник; 19 — жировое тело; 20, 21 — яйцеводы; 22 — маточный отдел яйцеводов; 23— отверстие яйцеводов в клоаке; 24 -спинная аорта; 25—задняя полая вена; 26— сонная артерия; 27 - дут аорты; 28-'- легочная артерия
В процессе эмбрионального развития легкое образуется как слепой вырост переднего (глоточного) отдела пищеварительной трубки. Поэтому и во взрослом состоянии легкие остаются связанными с глоткой.
Отношение общей площади поверхности легких к площади кожи у амфибий составляет 2 : 3 (сравните, у млекопитающих внутренняя поверхность легких в 50—100 раз больше поверхности кожи). Поэтому для амфибий огромное значение имеет дополнительный газообмен, в первую очередь — кожный. За счет кожного дыхания в организм амфибий поступает 15—55% кислорода. Кожное дыхание имеет большое функциональное значение в качестве приспособления, обеспечивающего длительное нахождение животного в воде, например во время спячки. В этом случае жизнедеятельность поддерживается только за счет кожного дыхания.
Частично газообмен осуществляется и через слизистую оболочку ротовой полости, богато снабженной кровеносными сосудами. На такой тип дыхания приходится 10—15% потребляемого кислорода.
Газообмен у личинок амфибий осуществляется через кожу и с помощью разветвленных наружных жабр, которые у большинства видов исчезают в ходе метаморфоза. Однако у сирен и протеев жабры сохраняются в течение всей жизни.
У американских безлегочных саламандр и у дальневосточного тритона легкие полностью атрофированы — газообмен у них целиком происходит через кожу и слизистую оболочку ротоглоточной полости
Кровеносная система. В| связи с переходом к легочному газообмену и редукцией жаберного аппарата у амфибий появляется второй (легочный) круг кровообращения. Наличие двух кругов кровообращения невозможно с двухкамерным (как у рыб) сердцем, поэтому у всех наземных позвоночных в сердце имеются два предсердия — в правое предсердие поступает венозная кровь по венам большого круга, а в левое — артериальная кровь по венам малого круга кровообращения. Каждый круг начинается в желудочке и заканчивается в предсердии. От желудочка отходят артерии, причем в малом круге они несут венозную кровь, а в большом — артериальную. В предсердие впадают вены: в малом круге они приносят артериальную кровь, а в большом — трехкамерное, оно состоит желудочка (рис. 88).

Рис. 88. Вскрытое сердце лягушки: I — левое предсердие, 2 — клапаны, закрывающие общее отверстие, ведущее из обоих предсердий в желудочек, 3 — желудочек, 4 — артериальный конус, 5 — правое предсердие, 6 — кожно-легочная артерия, 7—дуга аорты, 8 — общая сонная артерия, 9 — левый артериальный ствол, 10 — спиральный клапан артериального конуса
У низших амфибий (безногих и хвостатых) левое и правое предсердия разделены не полностью. У бесхвостых — перегородка между предсердиями полная.
К правому предсердию примыкает тонкостенная венозная пазуха, в которую впадают вены, несущие венозную кровь от всех частей тела. В левое предсердие впадают легочные вены с артериальной кровью. От правой стороны желудочка отходит артериальный конус, разделяющийся затем на правый и левый артериальные стволы. Каждый ствол, в свою очередь, разделен продольными перегородками на три сосуда: кожно-легочную артерию, дугу аорты и общую сонную артерию. Артериальный конус имеет спиральный клапан, распределяющий поступающую из желу дочка кровь в эти 3 пары сосудов. В упрощенном виде процесс распределения крови можно представить следующим образом. При сокращении желудочка первая порция наиболее венозной крови из правой части желудочка поступаем в первые отходящие сосуды — кожно-легочные артерии, по которым она доносится до органов дыхания (кожи и легких). Отверстия других сосудов в это время закрыты спиральным клапаном. При дальнейшем сокращении желудочка давление в его полости растет, при этом спиральный клапан смещается, и следующая порция смешанной крови из центральной части желудочка попадает в дуги аорты. В кожно-легочные артерии эта кровь не поступает, поскольку они уже заполнены кровью. Когда сокращение миокарда желудочка подходит к концу, еще более возросшее давление сдвигает спиральный клапан дальше, и артериальная кровь из левой части желудочка попадает в сонные артерии и по ним — в голову.
От дуг аорты отходят подключичные артерии, несущие кровь к передним конечностям. Описав полукруг и обогнув сердце, дуги аорты объединяются в общий сосуд — спинную аорту, от которой отходят кровеносные сосуды к внутренним органам и сосуды, снабжающие кровью задние конечности, — общие подвздошные артерии.
Каждая кожно-легочная артерия, содержащая преимущественно венозную кровь, разделяется на легочную артерию, несущую кровь в легкие, и кожную артерию, которая ветвится в коже спинной поверхности тела. После окисления в легких кровь становится артериальной, собирается в легочные вены и по ним впадает непосредственно в левое предсердие.
Венозная кровь от головы и передних конечностей собирается в две передние полые вены, которые получаются в результате слияния яремных и подключичных вен. Передние полые вены принимают в себя кожные вены, несущие артериальную кровь. Смешанная кровь по передним полым венам попадает в венозную пазуху.
Часть венозной крови от задних конечностей идет к почкам и проходит через воротную систему (сосуды распадаются на сеть капилляров). Выходящие из почек почечные вены сливаются, образуя непарную заднюю полую вену.Воротная вена печени, несущая кровь от органов пищеварительной системы, и брюшная вена с венозной кровью от задних конечностей и мочевого пузыря образуют воротную систему печени. Очищенная в печени венозная кровь собирается в две короткие печеночные вены, которые впадают и заднюю полую вену. В конечном итоге задняя полая вена, несущая кровь от туловища и задних конечностей, впадает и венозную пазуху.
Относительно простое строение кровеносной системы, снабжение органов смешанной кровью и слабо развитая дыхательная система обеспечивают у амфибий достаточно низкий уровень обмена веществ. Частота сокращений сердца у травяной лягушки составляет примерно 40—50 раз и минуту, что на порядок меньше по сравнению с птицами такой же массы. Поэтому у амфибий медленный кровоток и низкое давление крови — у бесхвостых около 30/20 (для сравнения: у чешуйчатых рептилий — 80/60). Эритроциты v амфибий — крупные ядерные клетки, число которых невелико (от 20 тыс. до 730 тыс. в 1 мм3 крови). У земноводных появляется новый кроветворный орган — красный костный мозг трубчатых костей.
Нервная система и органы чувств. Головной мозг 1емноводных по сравнению с рыбами имеет ряд прогрессивных особенностей. Довольно крупный передний мозг хорошо развит и разделен на полушария. У земноводных формируется свод мозга — первичная кора (архипаллиум), которая среди костных рыб присутствует только у двоякодышащих. Промежуточный мозг имеет эпифиз и хорошо развитый гипофиз. Средний мозг небольшой. Мозжечок развит слабо, что связано с простой двигательной активностью активностью земноводных. От головного мозга, как и у рыб, отходит 10 пар черепно-мозговых нервов.
В связи с выходом на сушу органы чувств земноводных претерпевают значительные изменения.
Органы обоняния представлены парными обонятельными капсулами, которые сообщаются с внешней средой через наружные ноздри, а с ротоглоточной полостью — через внутренние ноздри (хоаны). Органы обоняния функционируют только в воздушной среде, в воде ноздри закрыты.
Выпуклая роговица глаза и более плоский, чем у рыб, двояковыпуклый хрусталик позволяет различать предметы в воздушной среде на значительном расстоянии. Размер округлого или щелевидного зрачка может значительно изменяться в зависимости от интенсивности освещения. Появляются подвижные верхние и нижние веки, которые защищают глаз, а также третье веко, или мигательная перепонка, которая расположена в переднем углу глаза. Секрет специальных слезных желез обеспечивает влажность роговицы в сухой воздушной среде. Аккомодация зрения осуществляется путем смещения хрусталика вперед под действием специальной ресничной мышцы. По-видимому, некоторые амфибии обладают цветовым зрением: в сетчатке есть палочки и колбочки.
У амфибий с сумеречной активностью преобладают палочки. Число фоторецепторных клеток у хвостатых амфибий 30— 80 тыс. на 1 мм2 сетчатки, у бесхвостых значительно больше — 400—680 тыс.
Звук в воздушной среде распространяется гораздо хуже, чем в воде, поэтому орган слуха у земноводных организован гораздо сложнее, чем у рыб. Образуется новый отдел — среднее ухо, которое представляет собой небольшую полость в черепе (ее называют барабанной), заполненную воздухом и выстланную слизистой оболочкой. Снаружи барабанная полость прикрыта тонкой барабанной перепонкой, воспринимающей звуковые колебания. С ротовой полостью ее связывает слуховая (евстахиева) труба, которая служит для выравнивания давления между внешней средой и полостью среднего уха. Верхний элемент подъязычной дуги превращается у амфибий в слуховую косточку — стремечко, имеющую вид палочки, одним концом упирающейся в барабанную перепонку, а другим — в овальное окно, ведущее в полость внутреннего уха. Стремечко усиливает звуковые колебания воздушной среды, вызывая вибрацию эндолимфы в перепончатом лабиринте.
Данные сравнительной анатомии и эмбриологии показывают, что полость среднего уха у амфибий гомологична брызгальцу рыб, т. е. рудиментарной жаберной щели, лежащей между челюстной и подъязычной дугами.
Лягушки воспринимают звуки частотой от 30 до 15 тыс. Гц. Наличие голосовых связок позволяет многим бесхвостым амфибиям издавать звуки, которые усиливаются резонаторами.
У личинок амфибий имеются органы боковой линии, которые во взрослом состоянии сохраняются только у водных форм.
Ныделительная система. У взрослых амфибий выделительная система представлена парой первичных туловищных (мезонефрических) почек, которые располагаются но бокам крестцового отдела позвоночника. Выводными протоками (мочеточниками) служат вольфовы каналы, которые у самцов одновременно выполняют функцию и семяпроводов. Мочеточники открываются самостоятельными отверстиями в клоаке. Сюда же у высших амфибий открывается мочевой пузырь. Моча из клоаки поступает в мочевой пузырь, где временно накапливается. После наполнении мочевого пузыря моча поступает обратно в клоаку, откуда выводится наружу.
Через почки у амфибий удаляются продукты обмена и излишняя вода, которая поступает в тело животного через кожу. Основным продуктом белкового обмена у взрослых амфибий является мочевина, а у водных личинок — аммиак, который в виде раствора выводится через жабры и кожу.
Количество нефронов в почках зависит от того, насколько тесно жизнь животного связана с водой. У водных хвостатых амфибий в каждой почке находится около 200—250 нефронов, а у бесхвостых — около 1 тыс. Это связано с тем, что водные амфибии часть продуктов метаболизма выделяют через жабры и покровы тела в окружающую их воду. Следует также отметить, что нефроны почек хвостатых амфибий устроены более примитивно — они имеют воронки, открывающиеся в полость тела.
Половая система, размножение и развитие. У самок парные яичники имеют зернистую структуру, их размеры варьируют в зависимости от стадии репродуктивного цикла. Особенно велики яичники весной, через их тонкую стенку просвечивают крупные яйца, богатые желтком. Рядом с яичниками располагаются тонкие, длинные яйцеводы (мюллеровы
каналы) и многолопастные жировые тела —запас питательных веществ для развития яйцеклеток. Каждый яйцевод одним концом (воронкой) открывается в полость тела, другим — в клоаку.
Созревшие яйца через разрыв стенок яичника попадают н полость тела, подхватываются воронкой и двигаются по яйцеводу, покрываясь слизистыми оболочками, которые затем сильно разбухают в воде.
У самцов земноводных сбоку от почек располагаются парные бобовидные семенники с примыкающими к ним жировыми телами (рис. 89). Жировые тела служат источником питательных веществ при сперматогенезе. Идущие от семенников тонкие семявыносящие канальцы проходят через почку и впадают в вольфов канал. По нему половые продукты попадают в клоаку и, минуя мочевой пузырь, выводятся наружу. У самцов земноводных вольфов канал совмещает функцию мочеточника и семяпровода
Рис.
89.
Мочеполовая система самца лягушки: 1 —
семенники,
— семявыносящие
канальцы,
— почки,
4
— надпочечники, 5— мочеточник (он же
семяпровод),
6 —
семенной пузырек, 7 — мочеполовое
отверстие,
8
— полость клоаки,
9
— отверстие мочевого пузыря,
10
— мочевой пузырь,
11 —
жировое тело
Оплодотворение у амфибий обычно происходит в воде, у некоторых хвостатых и у безногих амфибий оплодотворение внутреннее. Развитие зародыша происходит в воде, с метаморфозом. Личинки земноводных по своей организации гораздо ближе к рыбам, чем к взрослым амфибиям. Они имеют наружные жабры, которые со временем заменяются внутренними, двухкамерное сердце, один круг кровообращения, орган боковой линии, хвост, который не только служит органом движения, но и выполняет дыхательную функцию. Питаются головастики в основном растительной пищей. В ходе метаморфоза у головастиков формируются сначала передние, а затем и задние конечности, развиваются легкие, исчезают жабры и преобразуется кровеносная система. Заканчивается формирование скелета, постепенно рассасывается хвост, и головастик превращается в миниатюрную копию взрослой особи.
У некоторых видов хвостатых амфибий встречается неотения — способность к половому размножению на стадии личинки. Это явление хорошо изучено у аксолотля, личинки американской тигровой амбистомы. Экспериментально у аксолотля можно вызвать метаморфоз, давая ему препараты, содержащие гормон щитовидной железы — тироксин.
У некоторых видов амфибий появились приспособления, позволяющие уменьшить связь с водой во время размножения. Например, самец европейской жабы-повитухи наматывает икру на задние лапы и сидит с ней в норке в земле, но перед вылуплением головастиков отправляется в водоем. Некоторые квакши откладывают икру между листьями, нависающими над водой: вылупившиеся головастики падают в водоем. У южноамериканской пипы развитие икры вплоть до превращения личинок в маленьких лягушат происходит в особых кожных ячейках, образующихся на спине самки. Тритоны откладывают одиночные яйца, одетые более или менее плотной оболочкой.
Многообразие амфибий. Класс земноводных делится па три отряда: Безногие, Хвостатые и Бесхвостые. Общее число видов современных амфибий превышает 4 тыс.
Отряд Безногие. Самая малочисленная группа примитивных специализированных амфибий (около 170 видов), внешне напоминающих крупных червей или змей. К отряду относятся червяги и рыбозмеи (рис. 90). Обитают только во влажных тропических областях Азии, Африки и Америки.
1 Висцеральный скелет {висцеральный череп) — скелет рта и переднего отдела кишечника (глотки) у позвоночных. Развивается в результате дифференцировки висцеральных дуг предков — скелетных образований, расположенных в перегородках между жаберными щелями. У высших позвоночных носит название «лицевой череп».
2