

14. Разобщение окисления и фосфорилирования. Физиологическое и патологическое значение.
При действии факторов, разобщающих окисление и фосфорилирование, теплопродукция увеличивается. Так, 2,4-α-динитрофенол, угнетая процессы фосфорилирования, усиливает свободное окисление со значительным освобождением энергии и гиперпродукцией тепла.
Разобщение окисления и фосфорилирования вызывают также дифтерийный токсин, культуры стафилококка.
Тироксин, также обладающий разобщающим действием, вызывает набухание митохондрий, увеличивает проницаемость митохондриальной мембраны. Часть субстратов цикла Кребса выходит в гиалоплазму. Окисление их ферментами гиалоплазмы при участии экстрамитохондриального НАД приводит к увеличению образования тепла в результате свободного окисления. Выработка АТФ в клетке уменьшается.
При разобщении окисления и фосфорилирования наблюдается несоответствие между данными прямой и непрямой калориметрии. При отравлении 2,4-а-динитрофенолом теплопродукция по калориметру может на 50—80% превышать рассчитанную по газообмену. При тиреотоксикозе теплопродукция по калориметру может превышать рассчитанную по газообмену на 20—40%.
15. Коэффициент р/о как показатель эффективности окислительного фосфорилирования.
Коэффициент окислительного фосфорилирования
Окисление молекулы NADH в ЦПЭ сопровождается образованием 3 молекул АТФ; электроны от FAD-зависимых дегидрогеназ поступают в ЦПЭ на KoQ, минуя первый пункт сопряжения. Поэтому образуются только 2 молекулы АТФ. Отношение количества фосфорной кислоты (Р), использованной на фосфорилирование АДФ, к атому кислорода (О), поглощённого в процессе дыхания, называют коэффициентом окислительного фосфорилирования и обозначают Р/О. Следовательно, для NADH Р/О = 3, для сукцината Р/О - 2. Эти величины отражают теоретический максимум синтеза АТФ, фактически эта величина меньше.
16. Отношение атф/адф как регулятор окислительного фосфорилирования. Дыхательный контроль.
Дыхательный контроль – это регуляция скорости переноса электронов по дыхательной цепи отношением АТФ / АДФ . Чем меньше это отношение, тем интенсивнее идет дыхание и активнее синтезируется АТФ. Если АТФ не используется, и его концентрация в клетке возрастает, то прекращается поток электронов к кислороду. Накопление АДФ увеличивает окисление субстратов и поглощение кислорода. Механизм дыхательного контроля характеризуется высокой точностью и имеет важное значение, так как в результате его действия скорость синтеза АТФ соответствует потребностям клетки в энергии. Запасов АТФ в клетке не существует. Относительные концентрации АТФ / АДФ в тканях изменяются в узких пределах, в то время как потребление энергии клеткой может изменяться в десятки раз. Американский биохимик Д.Чанс предложил рассматривать 5 состояний митохондрий, при которых скорость их дыхания ограничивается определенными факторами: 1. Недостаток SH2 и АДФ – скорость дыхания очень низкая. 2. Недостаток SH2 при наличии АДФ – скорость ограничена. 3. Есть SH2 и АДФ – дыхание очень активно (лимитируется только скоростью транспорта ионов через мембрану). 4. Недостаток АДФ при наличии SH2 – дыхание тормозится (состояние дыхательного контроля). 5. Недостаток кислорода, при наличии SH2 и АДФ – состояние анаэробиоза.
17. Гормоны и лекарственные вещества как разобщители (механизм их действия).
18. Оксигеназное окисление: механизм, ферменты, значение.
19. Пероксидазное окисление: механизм, ферменты, значение.
20. Перекисное окисление липидов: механизм, значение. Антиоксидантная защита клетки.
Стадии перекисного окисления липидов
1) Инициация: образование свободного радикала (L•)
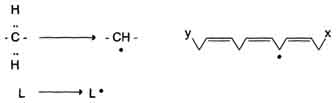
Инициирует реакцию чаще всего гидроксильный радикал, отнимающий водород от СН2-групп полиеновой кислоты, что приводит к образованию липидного радикала.
2) Развитие цепи:
L • + О2 → LOO • LOO• + LH → LOOM + LR•
Развитие цепи происходит при присоединении О2, в результате чего образуется липопе-роксирадикал LOO• или пероксид липида LOOH.
ПОЛ представляет собой свободнорадикальные цепные реакции, т.е. каждый образовавшийся радикал инициирует образование нескольких других.
3) Разрушение структуры липидов
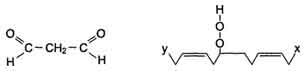
Конечные продукты перекисного окисления полиеновых кислот - малоновый диальдегид и гидропероксид кислоты.
4) Обрыв цепи - взаимодействие радикалов между собой:
LOO• + L• → LOOH + LH L• + vit E → LH + vit E• vit E• + L• → LH + vit Еокисл.
Развитие цепи может останавливаться при взаимодействии свободных радикалов между собой или при взаимодействии с различными антиоксидантами, например, витамином Е, который отдаёт электроны, превращаясь при этом в стабильную окисленную форму.