

6. Опишите механизм действия пиридинзависимых дегидрогеназ (механизм действия над в окислительно-восстановительных реакциях).
Механизм действия НАД и НАДФ. Коферменты НАД и НАДФ работают в клетке только вместе с белковой частью, образуя сложные ферменты дегидрогеназы. Специфичность связывания коферментов НАД и НАДФ с ферментами-белками зависит от адениннуклеотидной части их молекул, в то время как никотинамидная часть этих молекул осуществляет перенос электронов и протонов от окисляемого субстрата. Пиридиновое кольцо никотинамида имеет положительный заряд у четвертичного атома азота (поэтому окисленную форму НАД записывают в виде НАД+). Это вызывает смещение электронной плотности в пиридиновом кольце и появление положительного заряда у атома углерода в параположении. К нему присоединяется гидрид-ион (Н-), образующийся при неравномерном распределении электронов между атомами водорода: 2Н = Н- + Н+. Другой атом водорода, потерявший электрон (Н+), освобождается в окружающую среду, подкисляя ее. В результате образуется восстановленная форма НАД, записываемая в виде НАД.2Н или НАД.Н+Н+. Дегидрогеназные реакции с участием в качестве коферментов НАД и НАДФ имеют ряд характерных особенностей, которые обусловливают их ключевую роль в процессах биологического окисления. Первая особенность дегидрогеназных реакций - их легкая обратимость при небольших изменениях свободной энергии, что позволяет коферментам участвовать как в окислении субстрата, так и в восстановлении продуктов реакции (в зависимости от потребностей клетки). Вторая особенность заключается в способности этих коферментов (как в окисленной, так и в восстановленной форме) легко отделяться от белка-носителя, их высокой подвижности, что облегчает обмен атомами водорода и электронами между различными дегидрогеназными системами, расположенными в разных частях клетки. Коферменты НАД и НАДФ способны акцептировать водород от большого числа субстратов, окислительно-восстановительный потенциал которых ниже -0,3 В. К числу таких субстратов относятся продукты расщепления углеводов, жиров и различных аминокислот.
7. Опишите формулы окисленных и восстановленных форм флавиновых коферментов (ФМН, ФАД). Какая часть структуры ФМН и ФАД является акцептором (донором) в переносе электронов и протонов (формулы)? Частью каких комплексов они являются?
Флавиновые
ферменты –
это сложные ферменты, небелковая часть
которых представлена простетическими
группами ФМН (флавинаденинмононуклеотид)
или ФАД (флавинадениндинуклеотид).
ФМН
состоит из изоаллоксазинового кольца,
спирта рибитола (вместе они образуют
витамин В2 –
рибофлавин) и фосфорной кислоты, ФАД
содержит дополнительно адениловый
нуклеотид. Молекулярная структура ФАД
имеет вид:
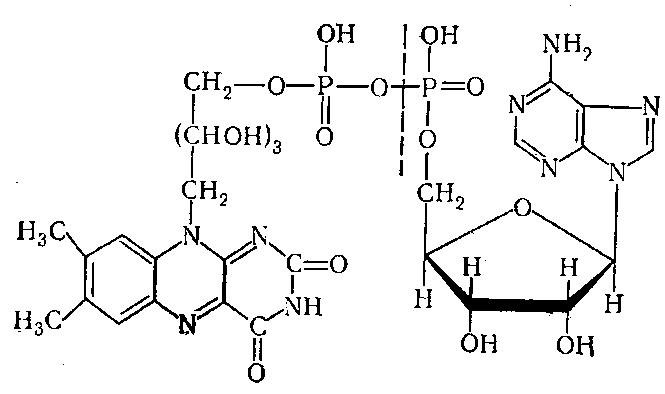 В
отличие от никотинамидных коферментов
ФМН и ФАД прочно (ковалентно) связываются
с соответствующими дегидрогеназными
белками и не отщепляются ни на одной из
стадий каталитического процесса.
Реакции, катализируемые флавинзависимыми
дегидрогеназами, трудно обратимы, и,
следовательно, флавиновые коферменты
не могут служить источником водородных
атомов в процессах
восстановительного биосинтеза, подобно
никотинамидным коферментам.
В
отличие от никотинамидных коферментов
ФМН и ФАД прочно (ковалентно) связываются
с соответствующими дегидрогеназными
белками и не отщепляются ни на одной из
стадий каталитического процесса.
Реакции, катализируемые флавинзависимыми
дегидрогеназами, трудно обратимы, и,
следовательно, флавиновые коферменты
не могут служить источником водородных
атомов в процессах
восстановительного биосинтеза, подобно
никотинамидным коферментам.