15. Чекпойнт перехода из g1 в s-фазу.
25. Митотический чекпойнт.
Концепция чекпойнтов — сверочных точек. Клетка решает идти ли дальше по клеточному циклу или лучше уйти в G0 / срочно исправить какие-нибудь косяки внутри себя и продолжить делиться / окончить свой земной путь апоптозом.
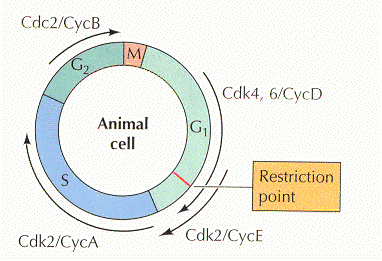
G1-S чекпойнт — клетка решает будет ли она делиться или уйдет на покой в G0. Участвуют белки:
CKI- p16 (CDK inhibitor p16) действует на CDK4/6, предотвращаяя взаимодейтсвие с D1. Активный комплекс CDK4/6-CYCLIN D фосфорилирует Rb, ответственный за ингибирование E2F. E2F вызывает экспрессию cyclin E, который взаимодействует с CDK2, позволяя начать дупликацию ДНК.
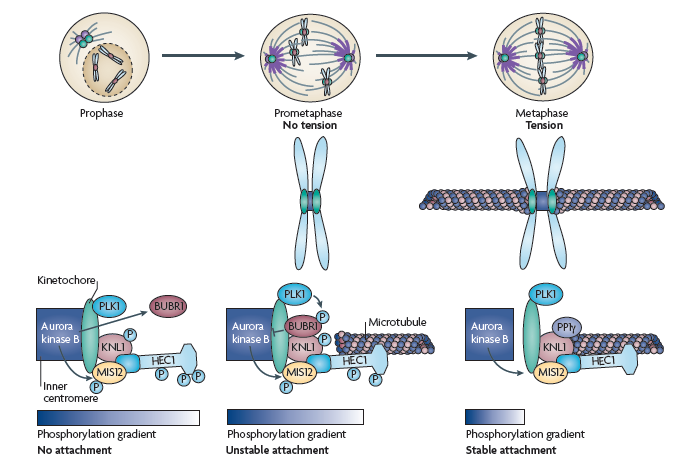
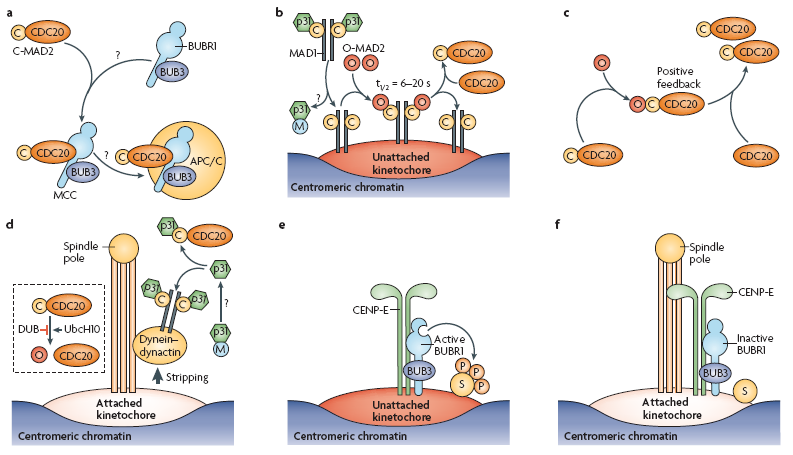
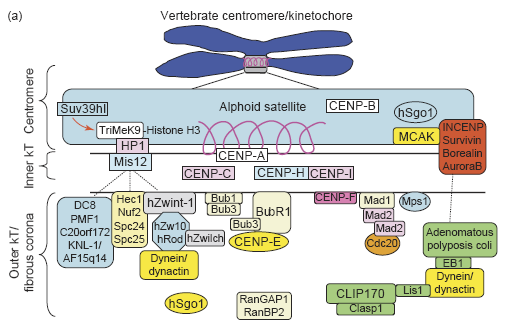
Митотический чекпойнт — проверяется все ли хромосомы соединены с мт. Если не все — то дальше пути нет! По-русски это называется spindle-assembly checkpoint (SAC). Как всегда, в этом деле замешан APC/C, на который действует cdc20, на который действует MCC. Участвуют белки: MAD (mitotic-arrest deficient) MAD1, MAD2, MAD3 (BUBR1 in humans) и многие другие гады (моторы, киназы, фосфатазы, и т. д.) Некоторые считают, что прохождение чекпойнта полностью зависит от фосфорилирования белков и создания градиента фосфорилирования с помощью Aurora, что имеет какие-то экспериментальные подтверждения
16. Возможные механизмы контроля размера клеток в клеточном цикле
1.Контроль скорости биосинтезов (прежде всего мониторинг трансляции)
2.Контроль массы или объема (например за счёт уровня фосфорилирования )
3.Контроль линейных размеров
Для метазоев (мухи, млеки) данный вопрос практически не изучен, были поставлены эксперименты на культурах клеток с текстурированными подложками, а также сгибающимися гелями, которые свидетельствуют о том, что клетка, оценив свою геометрическую конфигурацию либо пролиферирует, либо уходит в апоптоз. Как это происходит на молекулярном уровне в настоящее время неясно. Другой важный параметр – количество межклеточных контактов и/или контактов с субстратом, если их недостаточно клетка уходит в апоптоз.
Большинство работ по контролю размера проведены на дрожжах, однако глупо было бы экстарполировать эти результаты на ткани и культуры клеток.
Дробящиеся дрожжи содержат особый белок (Cdr2, расположен медиально), который при подготовке к митозу начинает измерять градиент других сигнальных белков (Pom1), который исходит от различных полюсов клетки. Данный контрольный механизм имеет место быть в S/G2, однако существуют и другие пока неизвестные механизмы.
19. Разнообразие Cdk
Комплекс CDK-циклин |
Позвоночные |
Почкующиеся дрожжи |
Делящиеся дрожжи |
|||
CDK |
циклин |
CDK |
циклин |
CDK |
циклин |
|
G1-CDK |
CDK 4,6 |
D (1, 2,3) |
CDK1= CDC28 |
CLN 3 |
CDK1= cdc2 (p34cdc2) |
Cln1, 2 DAF1 |
G1/S-CDK |
CDK2 |
E |
CLN1, 2 |
Cdc13 |
||
S-CDK |
CDK2 |
A |
CLB 5,6 |
Cig2 Clb5,6 |
||
M-CDK |
CDK1 |
B |
CLB 1,2, 3, 4 |
Cdc13 |
||
CDK1/cyclin B митоз
CDK2/cyclin A, E продвижение клеточного цикла
CDK3 G0/G1 progression, c-Jun phosphorylation
CDK4/cyclin D продвижение клеточного цикла
CDK5 регуляция цитоскелета, нейрогенез
CDK6/cyclin D продвижение клеточного цикла
CDK7/cyclinH часть общего транскрипционного фактора TFIIH,
CDK8/cyclinC остается с Mediator complex
CDK9/cyclinT формирует кор позитивного транскрипционного элонгпационного фактора b: P-TEFb.