

Глава 9
Внутриклеточные реакции на
изменения окружающей среды:
поиск оптимальной среды
существования
Резюме
В 1932 году Уолтер Кэннон, не обладая какими-либо специальными знаниями о метаболических и ферментативных процессах в клетке, написал книгу о мудрости тела, в которой использовал идеи французских и немецких физиологов конца XIX века. Адаптация - это ключевое понятие в биологии.Она бывает эволюционной, физиологической или культурной. Параметры оптимальной окружающей среды не могут быть постоянными, однако диапазон и частота их изменений должны иметь оптимальное значение.
Мудрость тела
В 1932 году произошло одно из величайших событий в биологии - Уолтер Кэннон опубликовал свою книгу под названием "Мудрость тела" [1], в которой нашли свое продолжение идеи его более ранней статьи "Организация физиологического го-меостаза" («Physiological Reviews», 1929) [2]. Незадолго до этого в своем научном докладе, представленном на праздновании юбилея Чарльза Ричета, Кэннон впервые использовал новый термин "гомеостаз". Однако это выступление, опубликованное в "Трудах Конгресса американских врачей и хирургов", осталось незамеченным и не получило такой известности, как его более поздние публикации.
В своей обзорной статье Кэннон приводит шесть основных постулатов, характеризующих гомеостаз. Так как только немногие из современных молекулярных биологов считали необходимым ссылаться на эти ранние, догматические представления о механизме обратной связи, я решил привести их в таблице 1.
129
ВАН РДНСЕЛЕР ПОТГЕР___________________________________________________________
Таблица 1. Шесть постулатов гомеостатической регуляции Уолтера Кэннона [2].
1. В открытой системе, которую представляет наш организм (состоящий из нестабильных компонентов, подверженных непрерывному разрушающему влиянию окружающей среды), сохранение постоянства само по себе есть свидетельство наличия сип, действующих или готовых действовать в направлении поддержания этого постоянства.
2. Сохранение постоянства объясняется тем, что любая попытка его нарушить автоматически встречает сопротивление в виде усиления действия одного или нескольких факторов.
3. Любой фактор, действие которого направлено на поддержание устойчивого состояния организма, не может одновременно работать в 2-ух противоположных направлениях.
4. Гомеостатические агенты, выступающие в качестве антагонистов в одной части организма, могут кооперировать — в другой.
5. Регулятивная система, определяющая состояние гомеостаза, может состоять из ряда взаимодействующих между собой факторов, вовлеченных в работу одновременно или последовательно .
6. Когда известна сила, которая может изменять гомеостатиче-ское состояние в каком-то одном направлении, следует искать механизм ее автоматического контроля или факторы, вызывающие прямо противоположный эффект.
Постулаты Кэннона, относящиеся к "мудрости тела", почти без особых изменений можно применить к новой группе явлений, под названием "мудрость клетки" — именно поэтому работы этого автора вызывает у нас такой интерес. Данная тема еще ждет своего развития (о чем можно судить на основании представленного ниже анализа книги Кэннона}. Однако уровень знания в период написания им книги, не позволял распространить шесть постулатов гомеостаза на "мудрость клетки". Насколько я могу судить, у Кэннона не было ни малейшего представления о ферментах в современном понимании этого слова, поэтому он даже не упоминает этот термин в своих работах. Для него понимание "активности клетки" включало все то, что сегодня изучается в виде схем и путей метаболизма, индукции, адаптации, репрессии, аллостерической активации и т. д. Не удивительно, что Кэннон, который и не помышлял о существовании каких-либо ферментов, не смог предвидеть развития приспособительных реакций за счет изменения количества фермента (хотя в метаболических процессах эти реакции встречаются достаточно часто).
130
________________________________________________БИОЭТИКА - МОСТ В БУДУЩЕЕ.
Вместе с тем, он в полной мере осознавал свой долг по отношению к предшественникам, в числе которых был великий французский физиолог Клод Бернар*. Бернар в 1878 году употребил знаменитое изречение о постоянстве milieu interieur* в таком контексте: "...все жизненные механизмы, при всем их разнообразии, имеют только одну цель — поддержание постоянными условий жизни во внутренней среде организма" (этот факт, безусловно, свидетельствует о вторичности концепции гомеостаза). Более того, еще в 1877 году известный немецкий физиолог Пфлюгер* независимо от Бернара утверждал: "Причина каждой потребности живого существа является также причиной удовлетворения этой потребности". В 1885 году Фредерик, а в 1900 году Шарль Рише* высказали похожую точку зрения. Так, Фредерик Рише пояснил: "Живое существо — это сила такого рода, которая при любом неблагоприятном воздействии сама по себе запускает компенсаторную реакцию по нейтрализации этого воздействия или восстановлению исходного состояния. Чем выше уровень развития живых существ, тем более разнообразными, совершенными и сложными становятся эти регуляторные механизмы. Они направлены на достижение полной [sic!] независимости от неблагоприятных факторов и изменений окружающей среды". Шарль Рише так говорил о живом организме: "В некотором смысле его состояние является стабильным благодаря его способности модифицироваться; легкая степень нестабильности — необходимое условие для достижения организмом подлинной устойчивости". Все ученые, на труды которых ссылается Кэннон, были знаменитыми физиологами. Эпоха биохимиков еще не настала.
Изменения Количества ферментов
В 1939 году началась новая эпоха. Блестящий молодой немецкий исследователь предложил концепцию метаболических циклов (за которую он позднее получил Нобелевскую премию) и стал известным как сэр Ганс Адольф Кребс из Оксфорда. Однако, факт существования цикла синтеза мочевины оспаривался на основании того, что экспериментальные данные были получены «из пробирки». Поэтому два американских ученых начали поиск дополнительных доказательств существования такого цикла. Они решили выяснить, увеличивается ли концентрация аргиназы, одного из ферментов орнитинового цикла, в печени крыс, если животных содержать на диете с высоким содержанием белка. Г. Лайтбоди и А. Клейнман [3J показали, что при этом активность фермента аргиназы действи-
внутренняя среда (фр.)
131
ВАН РАНСЕЛЕР ПОПЕР
тельно увеличивается, и таким образом получили дополнительные факты в пользу существования цикла Кребса и положили начало новой эре исследования ферментативных аспектов адаптации. При этом они провели различие между количеством и активностью ферментов: "В целом можно ожидать, что изменения активности ферментативных систем, вызванные разнообразием количества и качества пищи, могут приводить к развитию двух типов адаптации: (а) требующей принятия неотложных мер и (б) связанной с развитием медленно протекающих физиологических процессов. Приспособительные реакции первого рода включают изменения скорости и продолжительности действия ферментов, темпа поступления субстрата и удаления конечных продуктов реакции, активацию и инактивацию дополнительных количеств фермента. Второй тип адаптации может рассматриваться как адаптация путем изменения количества ферментов, необходимого для выполнения данной цели" (курсив авт.). Оказалось, что Кэннон не рассматривал механизм количественных изменений, так как все свое внимание он посвятил вопросу переключения функций органа от состояния покоя к состоянию работы и наоборот. Результаты ранних исследований Лайтбоди и Клейнмана в значительной степени были развиты в оригинальных экспериментах Р. Шимке и его соавторов [4], которые установили, что процессы синтеза и распада контролируются организмом независимо друг от друга.
Примерно в это же время методика определения количества ферментов в гомогенатах органов Поттера-Элевейема стала завоевывать широкое признание, и я опубликовал одну из моих ранних диаграмм, с изображением обратных связей между физиологией и молекулярной биологией [5]. Это было в 1944 году, когда такие понятия, как обратная связь и молекулярная биология, еще не появились в биохимической литературе. И, тем не менее, уже тогда я написал:
Специализированные ткани отличаются от более простых, растущих и неспециализированных, по крайней мере, тремя важными аспектами. Во-первых, они должны быть приспособлены для сохранения энергетических запасов до тех пор, пока не возникнет необходимость их быстрой мобилизации и использования для энергетического обеспечения определенной функции. Во-вторых, организм должен регулировать свои процессы таким образом, чтобы не препятствовать удовлетворению потребности функционирующей части. В-третьих, должен существовать механизм, при помощи которого специализированный орган сможет расти и развиваться для того, чтобы соответствовать тем потребностям, которые периодически испытывают его функциональные возможности. Первый аспект был изучен на примере активности ферментов и стадий промежуточного метаболизма. Второй - разработан физиологами, эндокринологами и фармакологами. Третий - представляет величайший интерес в связи с изучение раковых клеток, когда кажется, что их рост происходит, казалось бы, без всякой
132
____________________________________________БИОЭТИКА - МОСТ В БУДУЩЕЕ.
причины. Эти три этапа ответной реакции специализированной ткани на стимул являются, механизмами контроля, которые, в конечном счете, координируют свою работу (используя для этого общие знаменатели) в соответствии с действием естественных раздражителей. В данной статье в качестве такого общего знаменателя предлагается аденозинтрифосфатная система, которая обеспечивает интеграцию вышеназванных механизмов контроля ...".
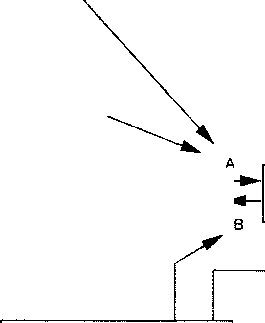
В своем обозрении покойный Джес Гринстейн так прокомментировал мою статью: "Ведущая роль АТФ в мышечном метаболизме признается давно. Но можно ли распространять данные, полученные при изучении мышечной физиологии, на другой вид тканей — этот вопрос, по-прежнему, остается открытым". [6] История последующих событий подтверждает возможность такой экстраполяции. В статье 1945 года (Рис. 9.1), посвященной исследованию проблемы шока и написанной мною совместно с В. Макшаном и соавторами [7], была вновь опубликована диаграмма 1944 года, отображающая характер обратных связей. Эта диаграмма представляет собой некий мост между работами Кэннона [1,2] и современными исследованиями по ферментативной природе клеточной адаптации.
Первой работой, получившей признание и открывшей веху изучения ферментативной адаптации у млекопитающих, был внушительный обзор В. Кнокса, В. Ауэрбаха и Э. Лина "Ферментативная и метаболическая адаптация животных" [8], который, также как и замечательная работа Кэннона, был опубликован в журнале "Physiological Reviews". Несмотря на достаточно большое количество цитируемых источников (их было 752), в исторической перспективе этот обзор оказался достаточно слабым. В нем, например, не было даже упомянуто имя Уолтера Б. Кэннона или понятие гомеостаза в связи с рассмотрением роли ферментативных приспособительных реакций, предназначенных для поддержания постоянства внутренней среды организма. Вместо того, чтобы обратиться к работе "Мудрость тела" авторы ссылались, как, впрочем, и их предшественники, на многочисленные работы бактериологов, в которых неоднократно утверждалось, что количество отдельных ферментов в бактериальной клетке может изменяться независимо от количества общего белка. Этот обзор стал героическим трудом, компиляцией различных источников для того, чтобы документально доказать факт существования ферментативной адаптации у животных. В нем практически не были рассмотрены связи описываемых фактов с физиологическими исследованиями в прошлом или теоретическими механизмами, недостаточно изучеными в настоящем. Такие механизмы в молекулярной биологии были рассмотрены В. Поттером и В. Ау-эрбахом в 1959 году [9], в преддверии бурных открытий в области исследования экспрессии гена как результата модифицирующего влияния окружающей среды.
133
ВАН РАНСЕЛЕР ПОТТЕР
Три Вида адаптации
Сегодня я могу сказать, что адаптация — это все то, что изучает современная биология. Если бы мне предложили прочитать только одну лекцию по биологии студентам, абсолютно не знакомым с этим предметом (что я фактически делаю один раз в год], я бы прочитал ее на тему адаптации.
От начала и до конца своих дней мы живем в условиях постоянно изменяющейся окружающей среды. И живем только потому что адаптируемся. Каждый новый год отличается от предыдущего, но то, чему мы научились сегодня, может пригодиться нам завтра. Если мы узнаем, что представляет собой процесс обучения, возможно, мы сможем учиться снова и снова.
Адаптация есть своего рода обучение и поэтому нам необходимо выяснить, в чем же она состоит. Если мы сможем понять, что адаптация — это путь к жизни, то, может быть, мы всерьез задумаемся над тем, в каком мире нам следует жить (если у нас еще есть выбор), и что является оптимальной окружающей средой для человечества? Именно эти вопросы я намерен обсуждать далее.
Первое, что мы должны уяснить - сегодня понятие "адаптация " не имеет четкого определения. Оно слишком перегружено в содержательном плане, поэтому только его точное определение может облегчить наше положение. Этот термин станет более ясным, если мы будем употреблять его вместе с определениями эволюционный, физиологический или куль турный.
Понятие "эволюционная адаптация" приложимо к жизни нескольких поколений популяции. При помощи эволюционной адаптации естественный отбор регулирует численность популяции, состоящей из неидентичных друг другу индивидов, и отбирает тех, чья наследственность в лучшей степени обеспечивает репродуктивную функцию в данной среде. Эволюционная адаптация не может предвидеть будущее, что часто (но не обязательно) приводит к исчезновению вида.
Мы нуждаемся в более глубоком понимании физиологической адаптации, поскольку приспособления такого рода, возникающие за считанные минуты или на протяжении многих недель и лет, присущи каждому из нас. По мнению У. Кэнно-на, физиологическая адаптация — это оркестр, исполняющий симсронию, состоящую из партий отдельных органов. И это действительно так. Но внутри органов обнаруживается мудрость отдельных клеток, или, так называемая, клеточная адаптация как механизм, в котором активность и количество ферментов то увеличиваются, то уменьшаются в зависимости от потребностей организма. Когда мы поднимаемся на большие высоты в горах, мы в определенной степени повышаем воз-
134
_________________________________________БИОЭТИКА - МОСТ В БУДУЩЕЕ.
можности наших клеточных механизмов; когда озабоченные своими делами, устремляемся по улице, вдыхая выхлопные газы, у нас активизируется другой набор ферментов (что будет показано далее).
Тканевая гипоксия
Повреждающее воздействие
"Шок" - цикл приводящий
к смерти
Аденозинтри фосфат (АТФ)
Уменьшение объёма циркулирующей крови
Геморрагия.
другие физиологические
факторы
Метаболические продукты
Адениловая Неорганический 20.000 кислота '• фосфат ~*~ кал.
Жизненные циклы
Гликолиз (молочная кислота) и дыхание (СС\)
Непосредсвенная
внутриклеточная
реакция
Увеличение потока крови
Глюкоза и кислород —
Г"
-*|— Вазодилатация -ч«— Локальная

Фармакологическая реакция ^.
ИН, ИНСУЛИН,
-неизвестные факторьГ
Увеличение синтеза биокатализаторов
Центральная
Развитие ответной реакции
Рис. 9.1.
Диаграмма обратных связей (1945 г.) АТФ выступает в качестве возможного общего знаменателя физиологических процессов в организме и его отдельных клетках при развитии ферментативной адаптации
(см. [7]).
135
ВАН РАНСЕЛЕР ПОПЕР
Наконец, культурная адаптация индивида является психологическим двойником физиологической адаптации, однако может быть аналогом и эволюционной адаптации, в процесс которой вовлечены целые популяции. Культурная адаптация, которую нам хотелось бы иметь представляет собой процесс распространения знаний обо всех видах приспособительных реакций организма и формирование более серьезного отношения к недостаточности наших знаний о тех аспектах адаптации, которые нам пригодятся для выживания в будущем. Более того, аналогия между шоком, вызванным влиянием культуры и физиологическим шоком позволяют нам оценивать эти явления как провалы в соответствующих видах адаптации.
Систематичес1ше 1солебания поведения Животных в условиях Контролируемого реЯшма питания
На протяжении последних лет в нашей лаборатории изучалась активность различных ферментов в печени белых крыс в связи с изучением природы перерождения нормальных клеток в раковые. Мы задали вопрос: "Аналогичны ли ответные ферментативные реакции нормальных и раковых клеток на воздействия одинаковых раздражителей?" По ходу работы стало ясно, что устойчивость определенных функций организма, показателями которой являются рН, щелочной резерв, температура тела и клеточный состав крови, при значительных изменениях внешней среды достигается за счет существенных сдвигов в активности ряда ферментов, концентрация которых то увеличивается, то уменьшается. Был начат эксперимент, согласно протоколу которого всех крыс разделили на три группы, получающие пищу, с содержанием 12, 30 и 60 % белка и 79, 61 и 31 % - углеводов, соответственно. Оставшиеся 9 % пищи составляли кукурузное масло, минеральные вещества и витамины [10J. Мы тщательно поддерживали световой режим: 12 часов, имитирующих день, и столько же часов, моделирующих ночное время суток. Причем животные имели доступ к пище только в течение первых 8 часов темного времени суток. Этот режим был обозначен нами как "8+16" (8 часов, когда пища доступна животным и 16 часов голодания). Для того, чтобы получить наиболее выраженные показатели изменения активности фермента и, отчасти для того, чтобы проверить нашу гипотезу о лучших показателях животных первой группы, вторая группа изучалась по схеме "8 + 40" (каждые два дня пища была доступна только первые 8 часов). Было доказано, что эти крысы весили в среднем на 100 г меньше, чем живот -
136
___________________ ___________________БИОЭТИКА - МОСТ В БУДУЩЕЕ.
ные "8+16" или же получавшие пищу ad libitum1 в течение месяца, (соответственно, 225-250 г - 1,11 и 325-350 г - 111). Недавно мы закончили эксперименты, длившиеся целый год с группой "8+16", в которой животным привили медленно растущие раковые опухоли. В течение всего периода эксперимента вес крыс оставался в пределах 250 г, и мы действительно убедились в том, что произошло значительное замедление роста опухоли. Полученные данные были опубликованы ("Cancer Research" 29:1691-98, 1969). В них было показано, что животные группы "8 + 40" могут достаточно долго жить и вести гиперактивный образ жизни (клетки имели оборудование для изучения двигательной активности крыс).
Во многих экспериментах, результаты которых приведены в литературе, одновременно измерялась активность только одного фермента. Появление частично автоматизированной аппаратуры сделало возможным увеличение количества измерений при помощи специальной методики профессора Х.С. Пи-тота. И теперь мы можем представить данные из последнего доклада В. Поттера, М. Ватанабе и X. Питота [10] об активности четырех ферментов у крыс, которые содержались в условиях трех различных режимов питания (Рис. 9.2). Во-первых, можно обнаружить, что активность тирозинтрансаминазы и сериндегидратазы увеличивается пропорционально содержанию белка в пище, однако синтез последней "не запускается" до тех пор, пока уровень белка не превысит 12 %. Во-вторых, активность глюкоза-6-фосфатдегидрогеназы и фермента, расщепляющего цитрат, увеличиваются пропорционально содержанию углеводов в пище и обратно пропорционально процентному содержанию белка. В-третьих, на уровень содержания глюкоза-6-фосфатдегидрогеназы влияет не только состав пищи, но также и длительность интервалов между кормлениями.
В разное время суток крысы периодически выводились из эксперимента, и было обнаружено, что в течение дня колеблется активность некоторых ферментов. Наиболее выраженными были изменения тирозинтрансаминазы, активность которой повышалась каждую ночь у животных в группе "8+ 16" [10J, а группе "8 + 40" ее активность повышалась дважды — ночью во время принятия пищи и вечером перед наступлением того дня, когда животные не получали пищи (Рис. 9.3., См.: [10]). Колебания активности тирозинтрансаминазы также наблюдались и в контрольной группе, однако, отклонения средних величин были менее выраженными [11]. В ходе экспери-
по желанию, по своему усмотрению (лат.). В данном случае имеется в виду контрольная группа животных, которой был предоставлен свободный доступ к пище в течение всего эксперимента (Прим. перев.).
137
ВАН РАНСЕЛЕР ПОТГЕР
600 i •. "8+16"
__ ______ БИОЭТИКА - МОСТ В БУДУЩЕЕ.
Рис. 9.2. Зависимость активности тирозинтрансаминазы, сериндегит-ратазы, глкжоза-6-фосфатдегидрогиназы и ферментов, расщепляющих цитраты в печени крыс, от содержания белка в пище. Заштрихованные фигуры, соединенные непрерывной, прерывистой и пунктирной линиями, соответствуют средним значениям указанных показателей у крыс из группы «8 + 40», «8+16» и «8+ 16». Первые две группы содержатся в обыкновенных клетках, последняя — в специально оборудованных для изучения двигательной активности клетках (См. [10])
мента по изучению способности печени к накоплению аналога аминокислоты было показано, что колебания активности фермента происходят только в опытных группах. Анализ данных, полученных в контрольной группе ad libitum, не выявил каких-либо систематических флуктуации. Эти разнообразные закономерности были недавно обобщены в материалах "Симпозиума по проблемам питания и метаболической регуляции" и поэтому нет необходимости их детализировать [11]. В указанном сообщении, мы привели пять определений различных целей, которые можно было бы использовать для исследования замены контролируемого режима питания животных режимом ad libitum с целью создания экспериментальных условий, релевантных состоянию благополучия человека. Одной из таких целей был поиск оптимального режима , питания с точки зрения различных критериев, таких как долголетие, фертильность, эффективность совершаемой работы. После симпозиума кто-то заметил, что указанные три критерия не рассматриваются нами на предмет "совместимости с другими критериями", и я согласился, что оптимальный режим питания должен учитывать не только те факторы, которые мы первоначально указали.
Flouck оптимальной оКруЖающей среды
Первоначально, разрабатывая режим "8 + 40", мы предполагали, что он обеспечит выживание животных, находящихся на низкокалорийной диете, а что состояние животных контрольной группы, получающих пищу по желанию, будет хуже, чем в естественных условиях. Имеется достаточно обширная литература (на которую мы частично ссылались [11]), где показано, что у животных, находящихся на ограниченном по калорийности режиме питания, спонтанные случаи развития опухолей случались реже и эти животные по ряду причин жили дольше, чем те, которые ели вдоволь. Нам кажется, что подопытные . животные, содержащиеся взаперти в маленькой клетке с обилием пищи под самым носом и которым нечего делать, кроме как все время есть, находятся в весьма неестественной ситуации, не соответствующей их природным инстинктам. Я полагаю, что здесь есть прямая аналогия с тем положением, в котором сегодня оказался человек, однако, я все же не хотел бы
139
ВАН РАНСЕЛЕР ПОПЕР
преувеличивать значение отдельного случая. Что касается крыс, то можно предположить, что естественный отбор, по-видимому, управлял процессом появления не только тех особей, которые каждую ночь испытывают потребность в еде, но и тех, которые могут выдержать отсутствие еды в течение какого-то времени. Исходя из этого, мы установили режим 8 + 40" и, как я упоминал ранее, мы получили данные по животным, находившимся на этом режиме более 13 месяцев. При этом, действительно, моделировались состояния переедания или голодания, но даже после окончания эксперимента мы не можем сказать, существуют ли какие-либо положительные стороны данного режима или же нет. В этой связи следует подчеркнуть, что ряд теоретических школ которые предлагают в качестве оптимального режима - питание частое и "небольшими порциями". По этому вопросу имеется много литературы, особенно хочется отметить работы К. Коха и соавторов [12, 13]. Большинство предыдущих экспериментов мы проводили, не используя режима "8 + 40", и понятно, что если подобная методика или ее модификация имеет какое-либо преимущество, это еще необходимо доказать. В настоящее время мы продолжаем эксперименты по этой методике и исследуем эффекты воздействия различных режимов питания на жировой метаболизм крыс. Предварительные результаты этой работы уже опубликованы ("Federation Proceedings" 29'1553-59 1970).
Что же касается исследования оптимальных условий окружающей среды, мы хотим возвратиться к нашим предыдущим рассуждениям и предупредить, что постоянство внешней среды, которое, в конечном счете, приводит к развитию определенных адаптивных способностей, абсолютно нежелательно для здоровья животного. Животное может адаптироваться не только к изменениям среды, но, так или иначе, рассчитывает на это (но не на любую степень ее изменений). Если мы принимаем эту философию, то можем сформулировать проблему оптимальной окружающей среды следующим образом: вместо того, чтобы спрашивать, что собой представляет оптимально постоянная окружающая среда, мы задаем себе вопрос, какова оптимальная частота и амплитуда изменений окружающей среды. А вместо выяснения, какова оптимальная температура среды, мы исследуем оптимальный ее диапазон и частоту изменений; и, наконец, - не ищем ответа на вопрос об оптимальной диете при постоянном наличии пищи, а задумываемся над тем, каковы оптимальные частота употребления и количество пищи.
При экстраполяции экспериментальных данных с животных на человека важно учитывать фактор совместимости его с другими людьми, то есть, результаты воздействия в социальной среде. Однако исследование такой совместимости с другими не является самоцелью. Мы скорее должны видеть проблему в
140
_________________________________________БИОЭТИКА - МОСТ В БУДУЩЕЕ.
перспективе, чем препятствовать поступлению жалоб в "Ящик жалоб и предложений". Наконец, мы должны спросить, какая же среда обитания будет в большей степени способствовать развитию и совершенствованию цивилизованного мира.
Активность тирозинтрансаминазы
Кормление
Голодание
' • * • «г"'';
![]()
600
500
i 400
I
> 300
200
100
" 0900 1500 2100 0300 0900 1500 2100
1200 1700 2400 0600 1100 1700
Время дня
Рис. 9.3.
Активность тирозинтрансаминазы в печени крыс (линия Holtzman), адаптированных к режиму «8 + 40» с содержанием белка в пище 12, 30 и 60%. Фигуры, соединённые непрерывной, прерывистой и пунктирной линиями, соответствуют средним значениям указанного показателя у крыс из групп с содержанием белка в пище — 12, 30 и 60%
(См. [10]).
Надо признать, что общество не предпринимало должных мер для необходимого контроля над степенью загрязнения окружающей среды. И только некоторые капризы развития цивилизации случайно привели к прозрению, что ряд инсектицидов в значительной степени снижает активность печеночных ферментов, выполняющих функцию детоксикации. Сегодня это уже ставший классическим, пример адаптации ферментов к изменениям окружающей среды. Но существует еще достаточно много соединений, которые индуцируют разнообразные ферменты-детоксиканты не только в печени, но в легких и кишечнике. Среди таких индукторов целый ряд веществ-транквилизаторов, обладающих седативными свойствами [14]. Напрашивается простой вывод, если мы можем производить вещества, обладающие ферментативной активностью, разру-
141
ВАН РАНСЕЛЕР ПОТТЕР
шающей токсические соединения окружающей нас среды, то зачем беспокоиться о ее загрязнении? Разве наши адаптивные способности не будут и далее совершенствоваться, и разве мы не сможем продолжать курить сигареты, дышать выхлопными газами двигателей и применять транквилизаторы? К сожалению, все это не так просто, как представляется некоторым. Мы слишком мало знаем обо всех возможных продуктах метаболизма лекарств. В действительности, некоторые соединения, которые сами по себе не являются канцерогенными, могут трансформироваться в соединения, вызывающие развитие рака [15]. Более того, любой вид адаптации следует рассматривать с точки зрения всеобщего равновесия, подразумевающего оценку всех ресурсов организма. В настоящий момент мы просто не можем сказать, существуют или нет какого-либо рода преимущества или недостатки для формирования адаптации при помощи, скажем, стократного увеличения активности фермента, который метаболизирует лекарство.
Заключение
Во-первых, я бы хотел подчеркнуть, что хотя сегодня достаточно много известно о генах, которые определяют потенциальную силу проявления любой способности организма, но все же нет гарантии, что такая способность обязательно проявится. Во-вторых, мы знаем, что экспрессия каждого гена регулируется как обратными, имеющими химическую природу связями с окружающей средой, так и результатами деятельности других генов в направлении формирования адаптации [16].
Для того, чтобы определить критерии оптимальной среды нам необходимо еще многое узнать, но уже сейчас ясно, что, нет такой среды, которая сама по себе могла бы считаться оптимальной. И если в духе философии плюрализма мы пытаемся найти ответ на вопрос об этих критериях, нам необходимо учитывать роль эволюционной, физиологической и культурной адаптации. Наконец, мы должны вспомнить высказанную мной прежде рекомендацию по поводу дальнейшего развития промышленности и сельского хозяйства: "Давайте будем использовать нашу потрясающую способность к производству для создания тех вещей, которые делают нас более мудрыми, а не более слабыми" [17]. Такие же мысли возникают у меня в связи с обсуждением феномена адаптации.
142
БИОЭТИКА - МОСТ В БУДУЩЕЕ.
ЛИТЕРАТУРА
1. Walter В. Cannon, The Wisdom of the Body (New York: W. W. Norton and Company, Inc., 1963).
2. Walter B. Cannon, "Organization for Physiological Homeostasis", Physiol. Reviews 9:399-431, 1929.
3. H. D. Lightbody, and A. Kleinman, "Variations Produced by Food Differences in the Concentration of Arginase in the Livers of White Rats", J. Biol. Chem. 129:71, 1939.
4. R. T. Schimke, "The Importance of Both Synthesis and Degradation in the Control of Arginase Levels in Rat Liver" J. Biol. Chem. 239:3808-3817, 1964.
5. V. R. Potter, "Biological Energy Transformation and the Cancer Problem", Advances in Enzymology 4:201-256, 1944.
6. J. G. Greenstein, book review, Cancer Research 5:62, 1945.
7. W. H. McShan, V. R. Potter, A. Goldman, E. G. Shipley, and R. K. Meyer, "Biological Energy Transformations During Shock as Shown by Blood Chemistry", Am J. Pbysiol. 145:93-106, 1945.
8. W. E. Knox, V. H. Auerbach and E. С. С. Lin, "Enzymatic and Metabolic Adaptations in Animals", Physiol. Review 36:164-254, 1956. (Contains 752 references).
9. V. R. Potter and V. H. Auerbach, "Adaptive Enzymes and Feedback Mechanisms", Lab. Invest. 8:495-509, 1959.
10. M. Watanabe, V. R. Potter and H. C. Pitot, "Systematic Oscillations in Tyrosine Transaminase and Other Metabolic Functions in Liver of Normal and Adrenalectomized Rats on Controlled Feeding Schedules", J. Nutr. 95:207-227, 1968.
11. Van R. Potter, E. F. Baril, M. Watanabe and E. D. Whittle, "Systematic Oscillations in Metabolic Functions in Liver from Rats Adapted to Controlled Feeding Schedules", Fed. Proc. 27:1238-45,
12. C. Cohn, "Feeding Patterns and Some Aspects of Cholesterol Metabolism", Fed. Proc. 23:76-81, 1964.
13. C. Cohn and D. Joseph, "Feeding Frequency and Upogenesis in Undernutrition", Can J. Pbysiol. Pharmacol. 45:609-612, 1967.
14. L. W. Wattenberg and J.L. Leong, "Effects of Phenothiazones on Protective Systems Against Polycyclic Hydrocarbons", Cancer Research 25:365-370, 1965.
15. E. C. Miller and J. A. Miller, "Mechanisms of Chemical Carcinogenesis: Nature of Proximate Carcinogens and Interactions with Macromolecules", Pharmacol. Reviews 18:805-838, 1966. (Contains 223 references.)
16. V. R. Potter, "Society and Science", Science 146:1018-1022, 1964. (Chapter 4, this volume, pp. 55-68).
17. V. R. Potter, "Bridge to the Future: The Concept of Human Progress", Land Economics 38:1-8, 1962. (Chapter 3, this volume, pp. 42-54).
143
ВАН РАНСЕЛ1Р ПОТПЕР