

12.3.2. Биосинтез ненасыщенных жирных кислот
Ненасыщенные жирные кислоты необходимы организму для синтеза полярных липидов, входящих в состав мембран, и нормального протекания обменных процессов.
Пальмитоолеиновая и олеиновая кислоты синтезируются из пальмитиновой и стеариновой кислот.
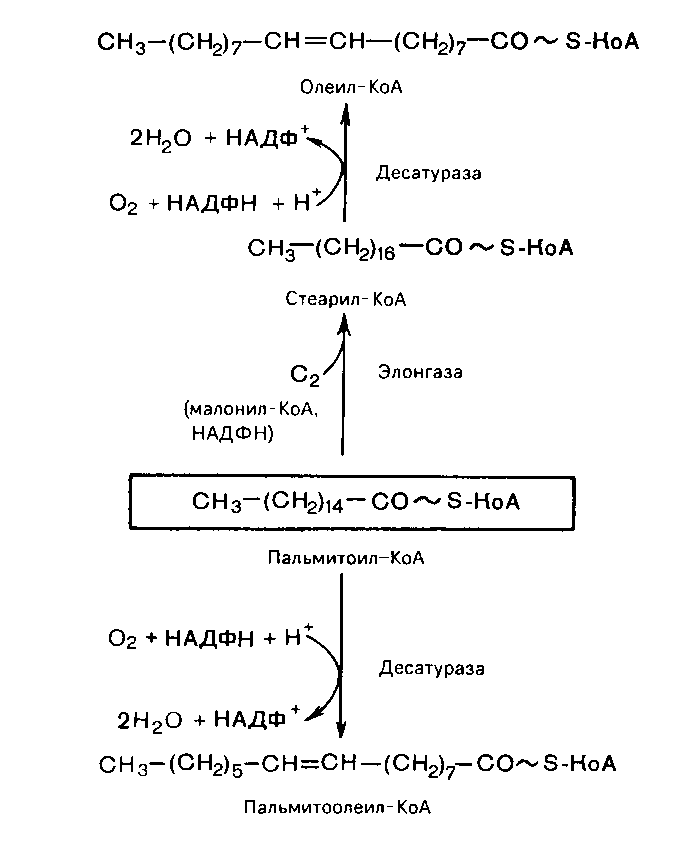
Эти превращения протекают в микросомах клеток печени и жировой ткани при участии молекулярного кислорода, восстановленной системы пиримидиновых нуклеотидов и цитохрома b5.
Однако в организме млекопитающих, в том числе и человека, не могут образовываться, например, линолевая и линоленовая кислоты. Они относятся к категории незаменимых и должны поступать с пищей. У растений насыщенные жирные кислоты превращаются в ненасыщенные с помощью дестуразной ферментной системы, которая вводит двойную связь в молекулу насыщенной кислоты независимо от ее углеродной цепочки.
12.3.3. Синтез триглицеридов
Организм синтезирует нейтральные жиры с целью пополнения запасов. Синтез триглицеридов происходит из глицерина и жирных кислот. Биосинтез триглицеридов протекает через образование глицерол-3-фосфата как промежуточного соединения. Глицерол-3-фосфат последовательно ацилируется двумя молекулами КоА-производного жирной кислоты, в результате образуется фосфатидная кислота (рис. 43).
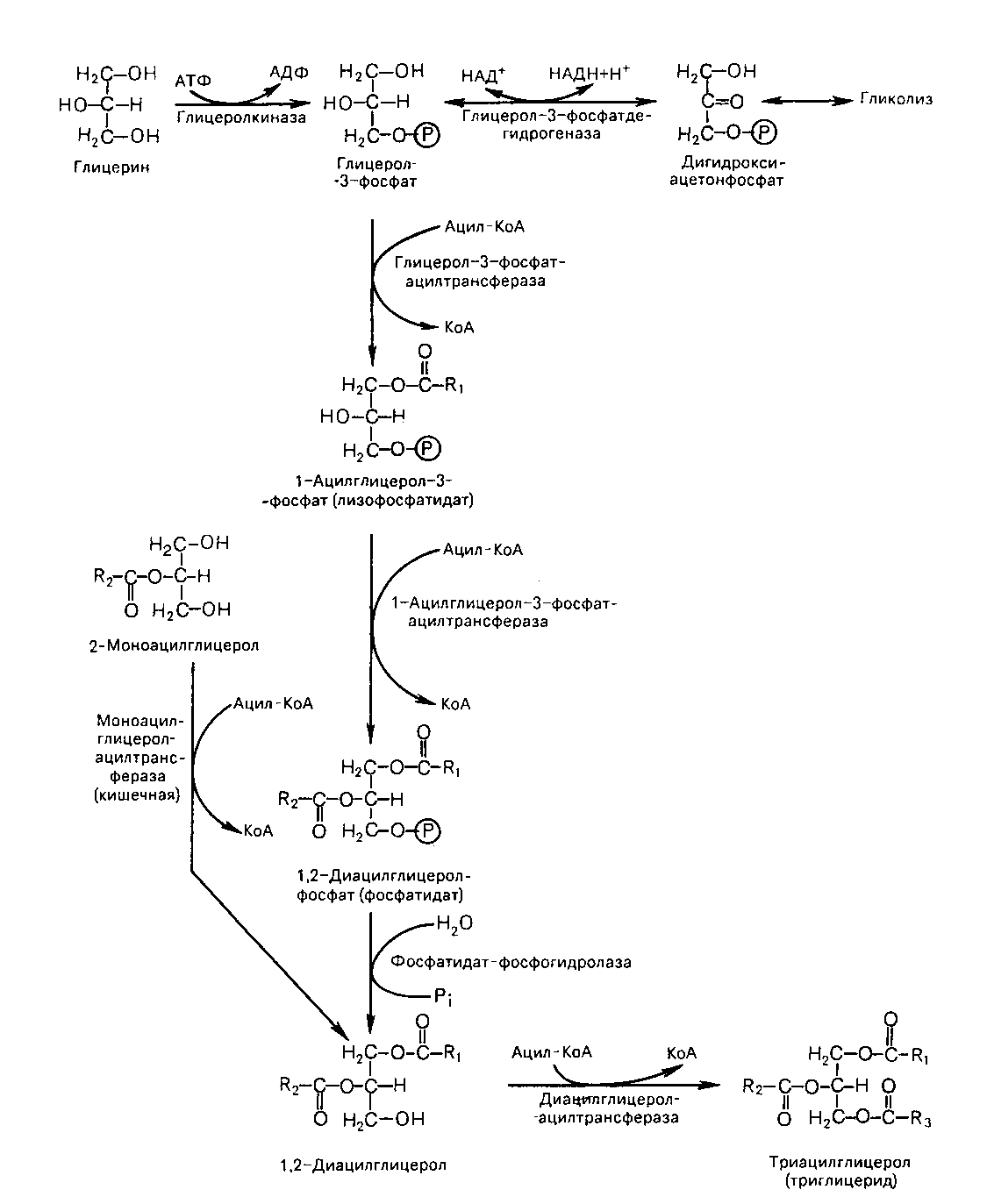
Рис. 43. Биосинтез триглицеридов
Далее фосфатидная кислота гидролизуется фосфатидат-фосфогидролазой до 1,2-диглицерида (1,2-диацилглицерола). Затем 1,2-диглицерид ацилируется третьей молекулой ацил-КоА и превращается в триглицерид (триацилглицерол).
Синтез глицерофосфолипидов.
Глицерофосфолипиды синтезируются во всех клетках (исключая зрелые эритроциты). Их активный синтез имеет место в стенке кишечника, печени, почках, мозге, мышечной ткани.
Фосфоглицерины различаются лишь природой спиртового остатка, связанного фосфоэфирной связью с фосфатидной кислотой. Для участия в синтезе фосфолипидов эти спирты предварительно «активируются» в ходе двустадийного процесса. На первой стадии они фосфорилируется с помощью АТФ:
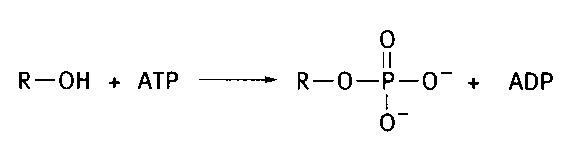
На второй стадии фосфорилированные спирты реагируют с цитидинтрифосфатом (ЦТФ аналог АТФ, в нем адениловый остаток заменен на цитозиновый):
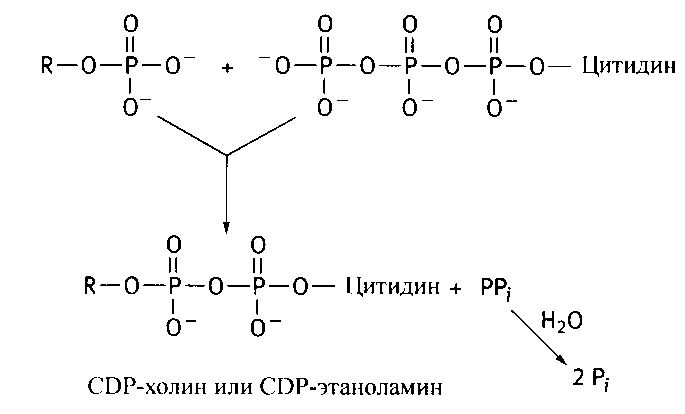
В конечной реакции синтеза фосфотидилэтаноламина и фосфотидилхолина участвует 1,2-диглицерид, образующийся под действием фосфатазы на фосфатидную кислоту:
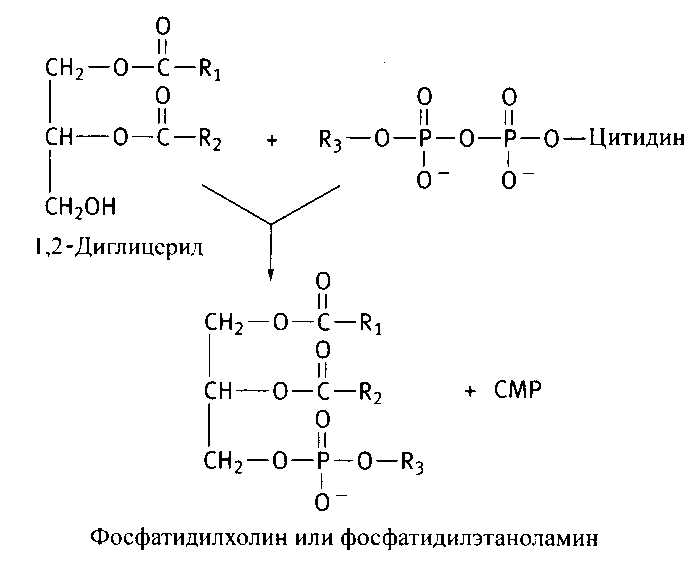
12.3.4. Биосинтез холестерина
Большинство клеток нашего организма (которые не утратили ядро) способны синтезировать холестерин. Печень и в меньшей степени тонкий кишечник – наиболее активные продуценты холестерина. Он является необходимым компонентом плазматических мембран и служит предшественником желчных кислот и стероидных гормонов.
Единственным исходным материалом служит ацетил-КоА. В синтезе холестерина выделяют три основные стадии: I – превращение активного ацетата в мевалоновую кислоту; II – образование сквалена из меваоновой кислоты; III циклизация сквалена в холестерин.
I стадия
Первая реакция синтеза холестерина представляет собой взаимодействие двух молекул ацетил-S-КоА под влиянием ацетоацетил-КоА-тиолазы, образуя ацетоацетил-S-КоА:
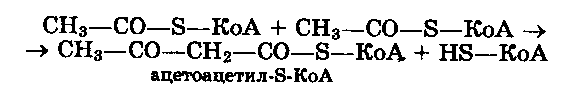
Полученный ацетоацетил-S-КоА конденсируется с третьей молекулой активного ацетата при участии специальной синтетазы, образуя -метил--гидрокси-глутарил-S-КоА:
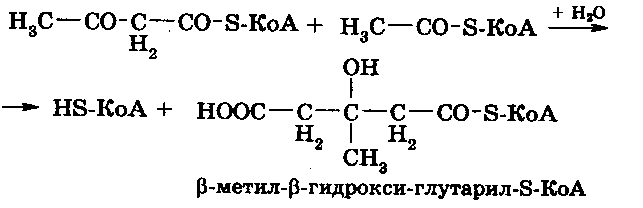
Следующая реакция восстановления -метил--гидрокси-глутарил-S-КоА под влиянием специальной редуктазы с образованием мевалоновой кислоты:
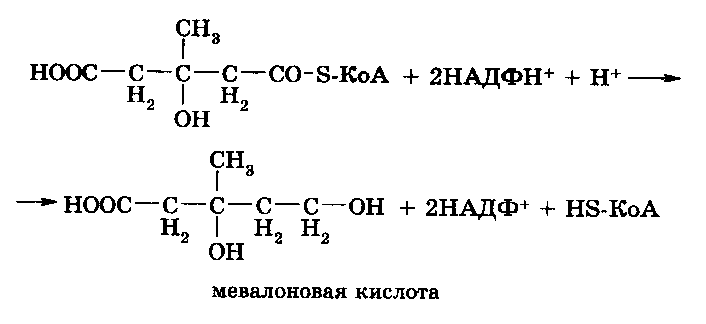
II стадия
Во второй стадии синтеза холестерола мевалоновая кислота превращается в сквален. Для этого мевалоновая кислота фосфорилируется с участием трех молекул АТФ, образуя 3-фосфо-5-пирофосфомевалоновую кислоту:
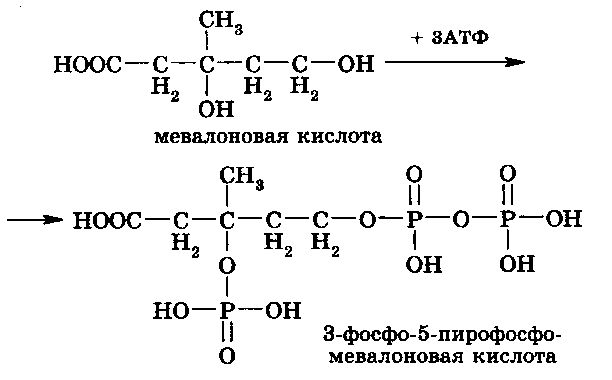
Нестабильная молекула 3-фосфо-5-пирофосфомевалоновой кислоты декарбоксилируется и дефосфорилируется, формируя непредельное соединение – изопентенил-пирофосфат (С5):

Изопентенил-пирофосфат изомеризуется, образуя 3,3-диметилаллил-пирофосфат(С5):
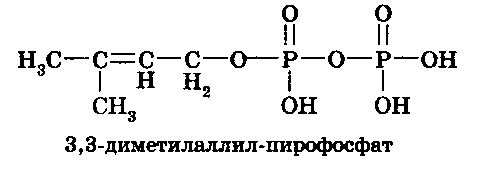
Конденсация двух 5-углеродных соединений (изопентенил-пирофосфат + 3,3-диметилаллил-пирофосфат) приводит к получению геранилпирофосфата (С5 + С5 = С10). Дальнейшая конденсация геранилпирофосфата с изопентенил-пирофосфатом (С10 + С5) дает образование фарнезил-пирофосфата (С15). В свою очередь, конденсация двух молекул фарнезил-пирофосфата заканчивается получением молекулы сквалена (С30).
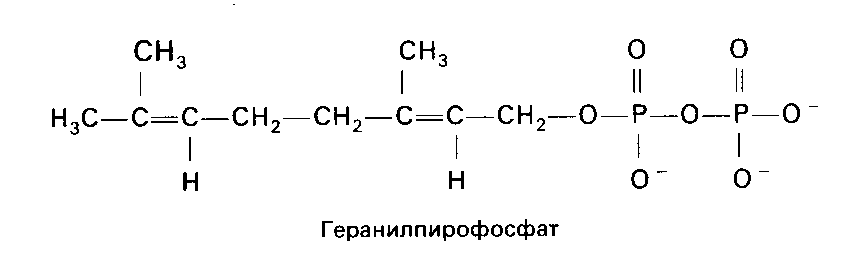

III стадия
На заключительной стадии следует циклизация сквалена при участии НАДФН+ + Н+, молекулярного кислорода и сквален-синтетазы с образованием ланостерина (С30), который уже имеет стероидную циклическую структуру. Ланостерин под влиянием ряда ферментов теряет три метильные группы и трансформируется в холестерол:
