

10.3. Обезвреживание аммиака в организме
Реакции дезаминирования аминокислот и азотистых оснований, окисление биогенных аминов приводят к образованию в тканях токсичного для организма аммиака. Аммиак токсичен и его содержание в крови должно быть предельно мало, иначе возможно нарушение функции мозга и развитие комы. В растениях аммиак содержится также в незначительных количествах. В живых организмах существуют механизмы обезвреживания аммиака.
Одним из путей связывания и обезвреживания аммиака является биосинтез глутамина (возможно аспарагина).
Основной механизм обезвреживания аммиака – биосинтез мочевины. Мочевина выводится с мочой; у растений может накапливаться в заметных количествах, особенно в растениях, имеющих на корнях микоризу.
Основным местом синтеза мочевины является печень. Образование мочевины происходит в результате нескольких ферментативных реакций, которые составляют орнитиновый цикл. Г.Кребс и К.Гензеляйт впервые в 1932 г. вывели уравнения реакций синтеза мочевины (рис.21).
Образование мочевины протекает в несколько этапов. На первом этапе синтезируется макроэргическое соединение карбомоилфосфат – метаболитически активная форма аммиака, используемая в качестве исходного продукта для синтеза пиримидиновых нуклеотидов и аргинина. В настоящее время открыты три разных пути синтеза карбомоилфосфата.
Первая реакция (необратимая) идет при участии фермента –аммиакзависимой карбомоилфосфатсинтетазы:
![]()
Реакция требует затраты двух молекул АТФ, открыта в митохондриях клеток печени и используется преимущественно для синтеза аргинина и мочевины.
Вторую, также необратимую, реакцию катализирует глутаминзависимая карбомоилфосфатсинтетаза:
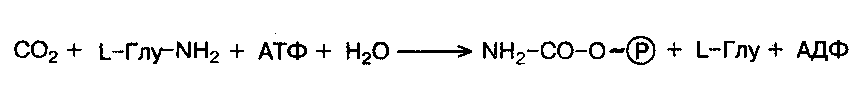
Реакция открыта в цитозоле клеток животных и требует наличия ионов Mg2+.
Третью обратимую реакцию катализирует карбаматкиназа (реакция открыта у микроорганизмов).
![]()
На втором этапе цикла мочевинообразования происходит конденсация кабомолфосфата и орнитина с образованием цитруллина; реакцию катализирует орнитин-карбомоилтрансфераза. На следующей стадии цитруллин превращается в аргинин в результате двух последующих реакций. Первая из них – конденсация цитруллина и аспарагиновой кислоты с образованием аргининосукцината (реакцию катализирует аргининосукцинатлиаза). Аргининосукцинат распадается в следующей реакции на аргинин и фумарат при участии аргининосукцинатлиазы. На последнем этапе аргинин расщепляется на мочевину и орнитин под действием аргиназы.
На синтез одной молекулы мочевины требуется затрата четырех высокоэнергетических фосфатных групп: две молекулы АТФ расходуются на синтез карбомоилфосфата и одна на образование аргининоянтарной кислоты, при этом АТФ расщепляется на АМФ и РРi, который при гидролизе также образует две молекулы Рi.
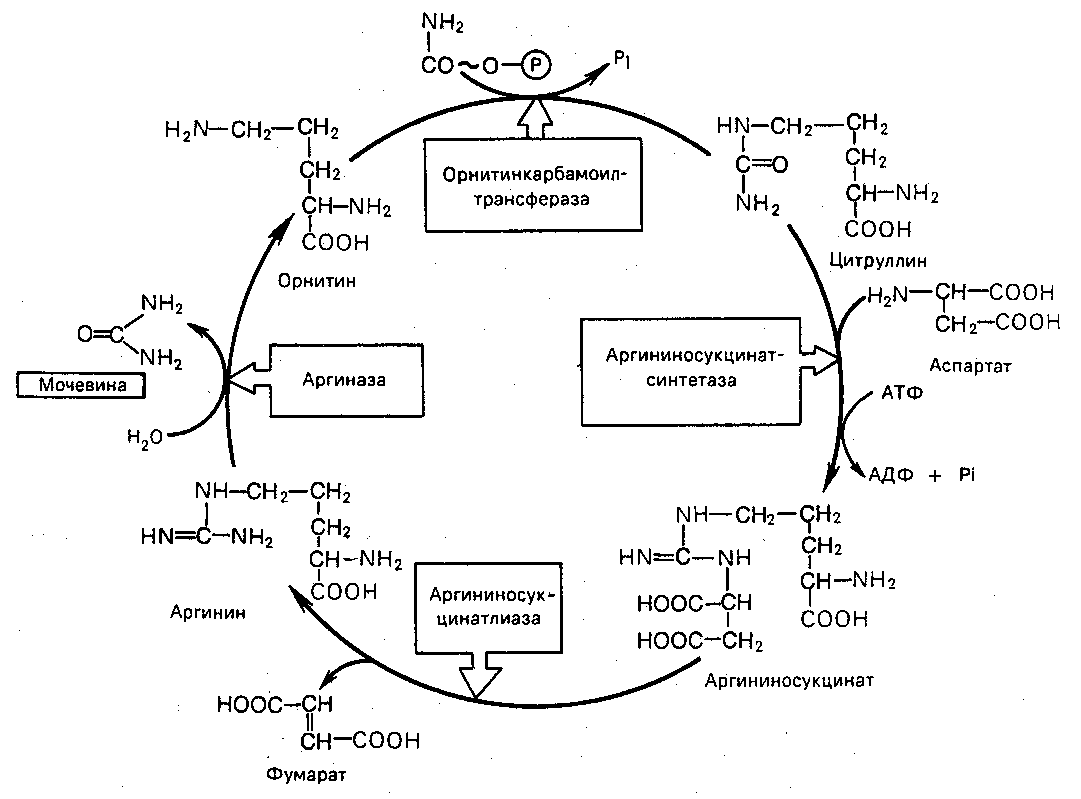
Рис. 21. Орнитиновый цикл синтеза мочевины в печени
В процессе эволюции живые организмы выработали различные типы азотистого обмена. Это аммониотелический тип, при котором главным конечным продуктом азотистого обмена является аммиак; он свойствен преимущественно рыбам. При уреотелическом типе обмена основным конечным продуктом обмена является мочевина; такой тип характерен для человека и животных. Урикотелический тип характерен для птиц и рептилий; главным конечным продуктом данного типа обмена является мочевая кислота.