

9.2. Синтез пиримидиновых нуклеотидов
Синтез пиримидиновых нуклеотидов начинается с элементарных уровней (СО2, NH3, аспартата).
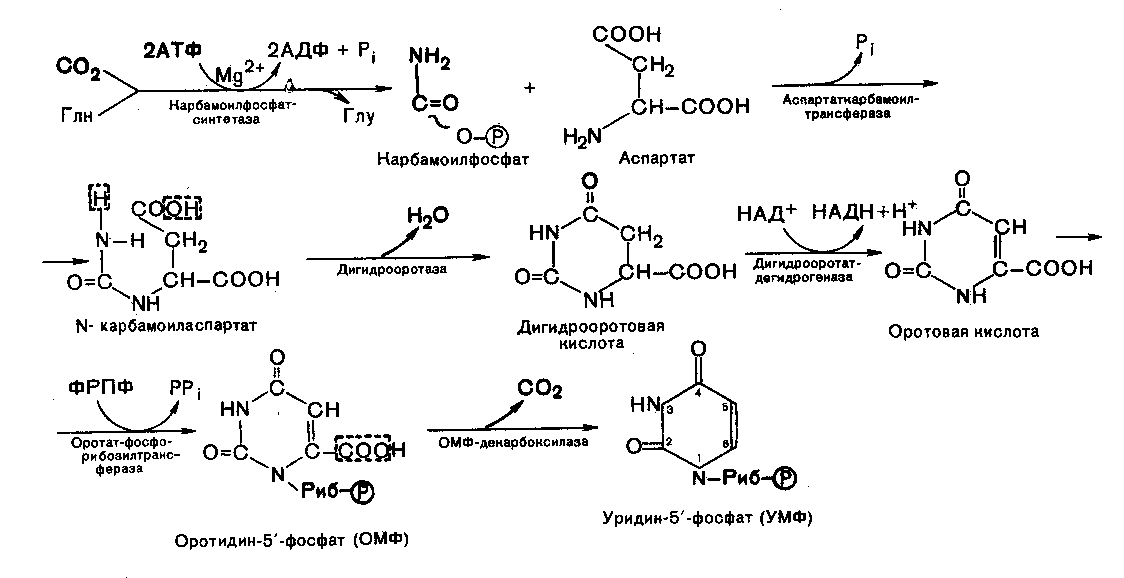
На первой стадии синтеза УМФ включает катализируемое цитоплазматической карбомоилфосфатминтетазой образование карбомоилфосфата из глутамина.
На второй стадии карбомоилфосфат реагирует с аспартатом, в результате чего образуется N-карбомоиласпарагиновая кислота. Последняя подвергается циклизации (под действием дигидрооротазы) с отщеплением молекулы воды, при этом образуется дигидрооротовая кислота, которая, подвергаясь дегидрированию, превращается в оротовую кислоту. В этой реакции участвует специфический НАД-содержащий фермент дигидрооротатдегидрогеназа. Оротовая кислота обратимо реагирует с ФРПФ, являющимся донатором рибозо-фосфата, с образованием оротидин-5/-фосфата (ОМФ). Дегидрокарбоксирилирование последнего приводит к образованию первого нуклеотида – уридин-5-фосфата (УМФ).
Превращение УМФ в УДФ и УТФ осуществляется, как и пуриновых нуклеотидов, путем фосфотрансферазных реакций:
УМФ + АТФ УДФ + АДФ
УДФ + АТФ УТФ + АДФ
УТФ может превращаться в цитидинфосфат (ЦТФ) за счет реакции аминирования с участием глутамина и фермента ЦТФ-синтетазы:
УТФ
+ Глн + АТФ
![]() ЦТФ + Глн + АДФ + Pi
ЦТФ + Глн + АДФ + Pi
Тимидиловые нуклеотиды входят в состав ДНК, содержащей дезоксирибозу. Превращение рибонуклеотидов в дезоксирибонуклеотиды сводится к восстановлению рибозы в 2-дезоксирибозу, требующему наличия двух атомов водорода. Источником последних является белок тиоредоксин, содержащий две свободные SH- группы на 108 аминокислотных остатков. Тиоредоксин легко окисляется, превращаясь в дисульфидную S-S-форму. Для его восстановления в системе имеется специфический ФАД-содержащий фермент тиоредоксинредуктаза, требующая наличия восстановленного НАДФН:
Тиоредоксин-(SH)2 + РДФ Тиоредоксин -S2 + dРДФ
Рибонуклеозиддифосфатредуктаза
Тиоредоксин-S2 + НАДФН+Н+ Тиоредоксин -(SН)2 + НАДФ+
Тиоредоксинредуктаза
При синтезе тимидинмонофосфата (ТМФ) требуется участие донора метильной группы, которым является N5, N10-метилен-ТГФК. В клетках имеется особый фермент тимидилатсинтетеза, катализирующая метилирование не свободного урацила, а dУМФ.

Синтез остальных дезоксирибонуклеозид-5/-трифосфатов, непосредственно участвующих в синтезе ДНК, осуществляется путем фосфорилирования дезоксирибонуклеозид-5/-дифосфатов в присутствии АТФ:
АТФ + dАДФ АДФ + dАТФ;
АТФ + dГДФ АДФ + dГТФ;
АТФ + dЦДФ АДФ + dЦТФ;
АТФ + dТДФ АДФ + dТТФ.
9.3. Биосинтез нуклеиновых кислот (биосинтез днк)
Во время деления клетки содержание ДНК должно удвоиться (реплицироваться). Репликация происходит полуконсервативным способом. Две исходные цепи, называемые родительскими, отделяются друг от друга: каждая служит матрицей для синтеза новой цепи, каждая новая двойная спираль содержит одну старую и одну новую цепь.
В основе образования новой цепи лежит принцип комплементарности оснований (Г с Ц и А с Т), так что последовательность оснований в родительской цепи однозначно определяет последовательность оснований в новой – дочерней – цепи.
Поскольку в ДНК пары оснований образуют водородные связи, синтез новых цепей может произойти только при расхождении цепей, что делает основания доступными для взаимодействия с комплементарными основаниями. Раскручивание или расплетание цепей осуществляется под действием фермента ДНК-геликазы. Цепи расходятся в определенной точке, называемой точкой начала репликации.
У прокариот образуются две репликативные вилки, которые движутся в противоположных направлениях. ДНК синтезируются со скоростью 500 пар оснований в секунду. Две вилки встречаются на противоположной стороне кольцевой хромосомы.
У эукариот синтез ДНК в репликативной вилке идет со скоростью 50 пар оснований в секунду, но начинается одновременно во многих точках (их, вероятно, более тысячи).
Цепи ДНК инициируются при помощи РНК. Специальная РНК-полимераза (праймаза) синтезирует небольшой участок РНК-праймера (затравочный олигонуклеотид, праймер) со свободной группой у С-3/ рибозы. Затем подключается ДНК-полимераза и продолжает наращивать цепь. Синтез ДНК всегда протекает в направлении 5/ 3/ растущей цепи. Это означает, что растущая цепь элонгирует (наращивается) в направлении 5/ 3/; нуклеотиды добавляются к свободному 3/-ОН-концу предшествующего нуклеотида. Синтез цепей в обратном направлении не происходит, поэтому синтезируемые цепи в каждой репликативной вилке должны расти в противоположных направлениях (рис. 19).
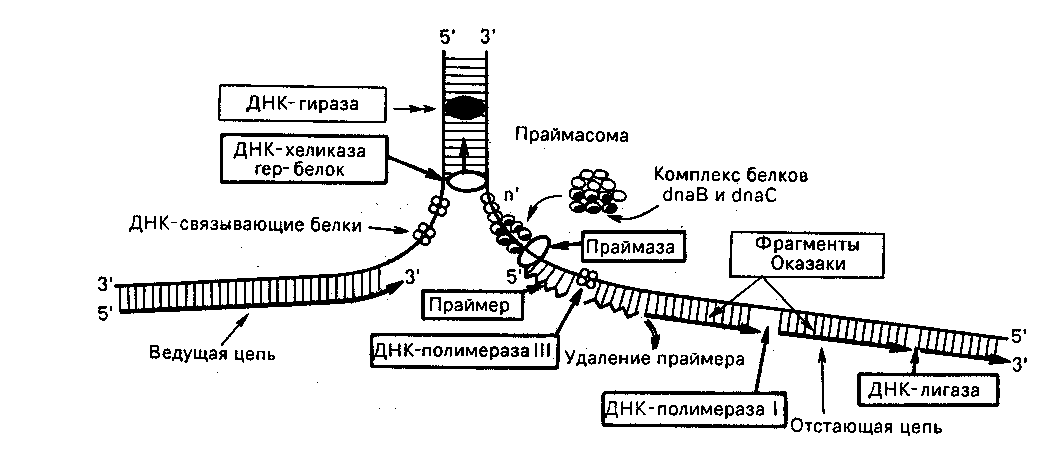
Рис. 19. Основные этапы репликации ДНК
При биосинтезе к цепям ДНК последовательно присоединяются ДНК-раскручивающие и ДНК-связывающие белки, а затем комплексы ДНК-полимераз и праймаз.
Синтез одной цепи (ведущей, лидирующей) происходит непрерывно, а другой (отстающей, запаздывающей) – импульсами (фрагментами Оказаки). Рост отстающей цепи идет то же от 5/ 3/, но в направлении, противоположном репликационной вилке. По мере движения репликативной вилки концы соседних фрагментов Оказаки соединяются при помощи ДНК-лигаз с образованием непрерывной отстающей цепи.
Этапы синтеза ДНК. Синтез ДНК может быть подразделен на три этапа: инициацию (начало синтеза), элонгацию (продолжение) и терминацию (завершение, прекращение синтеза). Каждый из этапов требует участия специфических ферментов и белковых факторов.
Инициация – первый этап в биосинтезе ДНК, является началом синтеза дочерних нуклеотидных цепей. В инициации участвуют минимум восемь различных ферментов и белков. Участок молекулы ДНК, в котором начинается репликация, называется репликатором, или областью начала репликации. Молекула ДНК, способная к автономной репликации, называется репликоном.
Раскручивание или расплетание цепей осуществляется под действием фермента ДНК-геликазы. В точке начала репликации образуется репликативная вилка.
Первоначально происходит ферментативный биосинтез на матрице ДНК необычного затравочного олигорибонуклеотида (праймера) со свободной группой у С-3/ рибозы. При инициации к цепям ДНК последовательно присоединяются ДКН-раскручивающие и ДНК-связывающие белки, а затем комплексы ДНК-полимераз и праймаз.
Элонгация – второй этап синтеза ДНК. Включает в себя синтез лидирующей и отстающей цепей. Синтез лидирующей цепи начинается у точки начала репликации. После синтеза праймера к нему присоединяются дезоксирибонуклеотиды под действием ДНК-полимеразы, далее синтез протекает непрерывно, следуя шагу репликационной вилки. Синтез отстающей цепи протекает в направлении, обратном движению репликационной вилки и начинается фрагментарно. Фрагменты Оказаки каждый раз синтезируются раздельно, начиная с синтеза праймера, который может переноситься с готового фрагмента при помощи одного из белковых факторов репликации в точку старта биосинтеза последующего фрагмента противоположно направлению синтеза фрагментов. Элонгация завершается отделением праймеров, объединением отдельных фрагментов ДНК при помощи ДНК-лигаз и формированием дочерней цепи ДНК.
Терминация – третий этап синтеза ДНК. Наступает, когда исчерпана ДНК-матрица и трансферазные реакции прекращаются. По завершении репликации образуются две молекулы двухспиральной ДНК, каждая из которых содержит одну материнскую и одну дочернюю, вновь синтезированную цепь. В результате митоза они поступают в дочерние клетки. Репликация обеспечивает воспроизведение генотипа в новых поколениях.