

Эфекторная функция ЦНС.
Ф-ии спинного мозга:
Моно- и полисинаптические рефлексы.
По степени сложности нейронной организации рефлекторных дуг различают моносинаптические рефлексы, дуги которых состоят из афферентного и эфферентного нейронов (например, коленный рефлекс), и полисинаптические рефлексы, дуги которых содержат также 1 или несколько промежуточных нейронов и имеют 2 или несколько синаптических переключений (например, флексорный рефлекс).
Клинически важные сухожильные рефлексы:
Коленный рефлекс, или пателлярный рефлекс — это безусловный рефлекс, относящийся к группе рефлексов растяжения, или стретч-рефлексов. Коленный рефлекс возникает при непродолжительном растяжении четырёхглавой мышцы бедра, вызванном лёгким ударом по сухожилию этой мышцы под надколенником. При ударе сухожилие растягивается, действуя в свою очередь на мышцу-разгибатель, что вызывает непроизвольное разгибание голени. Коленный рефлекс является классическим примером моносинаптического рефлекса.
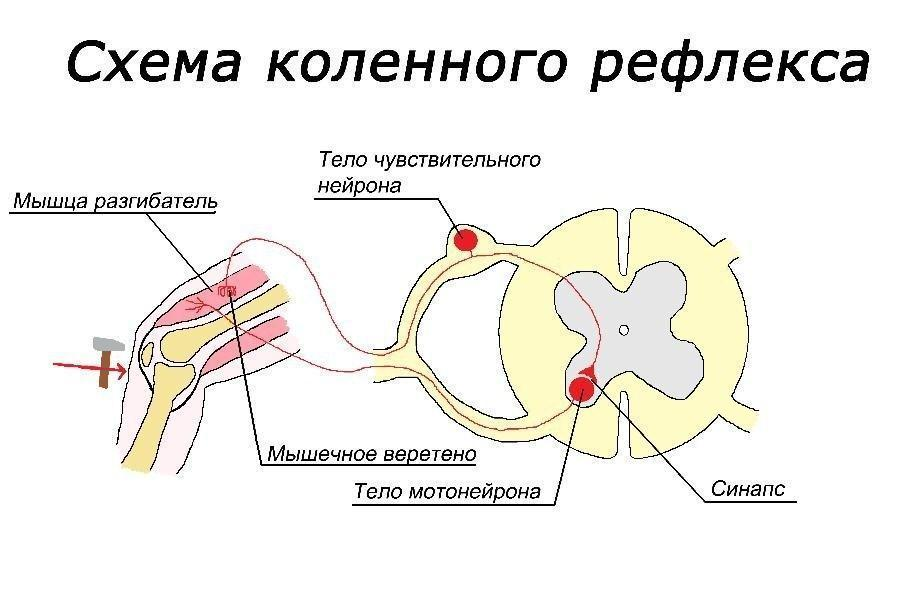
Сгибательно-локтевой, или рефлекс с сухожилия m. bicipitis, вызывается ударом молоточка по сухожилию двуглавой мышцы в локтевом сгибе. Ответной реакцией является сокращение названной мышцы и сгибание в локтевом суставе. Рефлекторная дуга: n. musculo-cutaneus(кожно-мышечный нерв), V и VI шейные сегменты спинного мозга. Глубокий, сухожильный рефлекс.
Рефлекс с трехглавой мышцы плеча (трицепс-рефлекс, разгибательно-лок-тевой рефлекс) вызывается ударом молоточка по сухожилию мышцы над локтевым отростком, при этом рука больного пассивно слегка отводится назад и кнаружи, предплечье в таком случае свободно свисает. Ответная реакция — разгибание предплечья. Афферентная и эфферентная части рефлекторной дуги проходят через локтевой нерв. Замыкается рефлекторная дуга в C7—С8 сегментах спинного мозга.
Защитные рефлексы.
Защитным рефлексом называют непроизвольное тоническое движение парализованной конечности в ответ на то или иное раздражение ее кожи. Защитные рефлексы у большинства больных обнаруживаются на нижних конечностях при поражении спинного мозга в его грудном отделе и чаще носят сгибательный характер.
Рефлекс сгибательного типа выражается непроизвольным сгибанием конечности в голеностопном, коленном и тазобедренном суставах, ведущим к «укорочению» конечности. Разгибательный рефлекс, наоборот, вызывает «удлинение» конечности вследствие непроизвольного выпрямления ее в тазобедренном, коленном суставах и подошвенного сгибания стопы.
Защитные рефлексы (чихание, кашель): Слизистая дыхательных путей просто усеяна рецепторами нервных окончаний, которые анализируют все происходящее в дыхательных путях. При попадании различных инородных тел и раздражающих веществ на слизистую оболочку дыхательных путей , а также при ее воспалении организм отвечает защитными рефлексами – чиханием и кашлем. Чихание возникает при раздражении рецепторов слизистой оболочки полости носа и представляет собой резкий выдох через нос, направленный на удаление раздражителя со слизистой. Кашель же является более сложным актом. Для того чтобы его произвести человеку необходимо глубоко вдохнуть, задержать дыхание, а затем совершить резкий выдох, при этом голосовая щель зачастую оказывается закрытой, что приводит к характерному звуку. Кашель возникает при раздражении слизистой гортани, трахее и бронхов. Основная задача защитных удаление раздражающих объектов с поверхности слизистых оболочек.
Защитный рефлекс новорождённого: Если новорождённого положить на живот, то происходит рефлекторный поворот головы в сторону. Этот рефлекс выражен с первых часов жизни. У детей с поражением ЦНС защитный рефлекс может отсутствовать, и, если не повернуть пассивно голову ребёнка в сторону, он может задохнуться.
Миотатический рефлекс растяжения.
Простейшим рефлексом яв-ся миотатический рефлекс, или рефлекс растяжения мышцы. Этот рефлекс можно вызвать у любой скелетной мышцы, хотя самый известный его пример — коленный рефлекс. Анат. основу миотатического рефлекса составляет моносинаптическая (с одним синапсом) рефлекторная дуга. Она включает сенсорный концевой орган, сенсорное нервное волокно с его клеточным телом в ганглии заднего корешка, α-мотонейрон, на к-ром сенсорный аксон образует синапс, и аксон этого α-мотонейрона, возвращающийся к мышце, от которой приходит сенсорное волокно. Сенсорным концевым органом в рефлексе растяжения мышцы служит мышечное веретено. Мышечное веретено имеет мышечные окончания, называемые интрафузальными волокнами, и центральную, немышечную область, связанную с окончанием афферентного нерва. Интрафузальные волокна иннервируются γ-мотонейронами передних корешков спинного мозга. Высшие центры головного мозга могут оказывать влияние на рефлекс растяжения мышцы посредством модуляции активности γ-мотонейронов. Этот рефлекс вызывается растяжением мышцы, что приводит к увеличению длины мышечного веретена и, вследствие этого, к повышению частоты генерирования потенциала действия в сенсорном (афферентном) нервном волокне. Повышенная активность в афферентном волокне усиливает разряд целевого γ-мотонейрона, что вызывает сокращение экстрафузальных волокон мышцы, от к-рой приходит афферентный сигнал. Когда экстрафузальные волокна сокращаются, мышца укорачивается и активность в афферентных волокнах понижается.
Висцеромотрные и вегетативные рефлексы спинного мозга:
Висцеромоторные рефлексы направлены на поддержание постоянства внутренней среды организма. Афферентная часть рефлекторных дуг этих рефлексов – сенсорные волокна черепно-мозговых нервов (зрительный, тройничный, лицевой, вестибулярный, слуховой, блуждающий). Эфферентная часть – аксоны мотонейронов ядер черепно-мозговых нервов либо нейроны спинного мозга и вегетативных ганглиев. Висцеромоторные рефлексы имеют четкие афферентные входы, легко возникают в процессе развития различных сложных защитных, приспособительных, пищевых и других рефлекторных реакций. На-пример, слезный рефлекс. Афферентная часть его рефлекторной дуги – сенсорные волокна слезной ветви тройничного нерва, распределенные в роговице. Этот рефлекс может возникнуть при подсыхании роговицы, при чихании, зевании, жевании, кашле, рвоте и даже при ярком освещении. Большая группа висцеромоторных рефлексов осуществляется через систему языкоглоточного и блуждающего нервов.
Спинальные вегетативные рефлексы — это замыкающиеся на уровне спинного мозга регуляторные реакции, осуществляющиеся при участии вегетативной нервной системы. В норме эти рефлексы находятся под жестким контролем со стороны вышележащих отделов центральной нервной системы. Перерыв спинного мозга приводит к развитию спинального шока — временному исчезновению соматических и вегетативных рефлексов. Чем выше уровень организации животного, тем больше длительность спинального шока: от нескольких минут у лягушки до нескольких месяцев у человека. Со временем явления спинального шока исчезают благодаря тому, что спинной мозг, отделенный от головного, начинает выполнять ряд регуляторных функций: восстанавливается регуляция сосудистого тонуса, рефлексы опорожнения мочевого пузыря и толстого кишечника, половые рефлексы.
Регулировать вегетативные функции за счет влияния на активность преганглионарных симпатических и парасимпатических нейронов могут практически все отделы центральной нервной системы, вплоть до коры больших полушарий. Наиболее тесно вегетативная нервная система связана со структурами, расположенными в стволе мозга и в гипоталамусе.
Проводниковые ф-ии спинного мозга:
Проводящие пути-отростки нервных клеток, соединяющие головной мозг со спинным. 4 типа: Восходящие(чувствительные, от различных рецепторов);
Нисходящие(двигательные, от коры к мотонейронам спинного мозга);
Комиссуральные(соединяют правую и левую половины мозга);
Ассоциативные(соединяют внутри половины мозга).
Восходящие пути.: Парные, перекрещенные.
-спинно-мозжечковые(передний и задний или пути Флексига и Говерса). К мозжечку поступает ин-ия от мышечных рецепторов.
-спинно-таламические(боковой и передний). Сигналы идут от тактильных рецепторов.
-пути задних канатиков или пути Голля и Бурдаха(нежный и клиновидный). Заканчиваются в теменной доле в постцентральной извилине, нач-ся в рец-орах.
Нисходящие пути:
-пирамидные(от коры).Делятся на передний(кортико-спинальный) и боковой(кортико-ядерный). Полностью перекрещенные.
-экстрапирамидные(от подкорковых центров). Делятся на красно-ядерно-спинальный, вестибулоспинальный, ретикуло-спинальные пути(латеральный и медиальный). Осознанные и рефлекторные движения.
Эффекторные функции ствола мозга.
В акте глотания различают три фазы:
Первая - оральная; твердая и жидкая пища при помощи языка направляется к заднему отделу ротоглотки.
Вторая - глоточная; рефлекс глотания вызывается касанием пищи задней стенки глотки. Регулируется центром глотания, находящимся в продолговатом мозгу.
Третья — пищеводная, она связана с координацией перистальтики пищевода и желудка. При глотании опущенные небные занавески поднимаются вверх и закрывают верхний отдел глотки, надгортанник временно закрывает вход в гортань. Пища поступает в пищевод, соединяющий глотку и желудок. Перед пищеводом находится трахея, за пищеводом - позвоночник. В пищеводе имеются три физиологических сужения. Первое - в месте перехода глотки в пищевод, второе - на уровне дуги аорты, третье - в месте перехода пищевода через отверстие диафрагмы. Продвижение пищевого комка по пищеводу происходит за счет волнообразных движений его стенок вследствие сокращения их мускулатуры.
Жевание — физиологический акт, заключающийся в измельчении с помощью зубов пищевых веществ и формировании пищевого комка. Жевание обеспечивает качество механической обработки пищи и определяет время ее пребывания в полости рта, оказывает рефлекторное возбуждающее влияние на секреторную и моторную деятельность желудка и кишечника. В жевании участвуют верхняя и нижняя челюсти, жевательная и мимическая мускулатура лица, язык, мягкое небо.
Механическая обработка пищи между верхними и нижними рядами зубов осуществляется благодаря перемещению нижней челюсти относительно верхней. У взрослого человека в ряду справа и слева имеются зубы разного функционального назначения — два резца и один клык (откусывающие пищу), два малых и три больших коренных, которые раздавливают и растирают пищу, — всего 32 зуба.
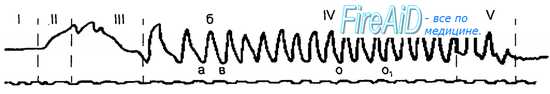
Рис. 11.10. Кимограмма жевательного периода. I — покой жевательной мускулатуры; II — фаза введения пищи в рот; III — ориентировочная фаза; IV — основная фаза; V — фаза формирования пищевого комка; а—б — опускание нижней челюсти, б—в — подъем нижней челюсти; о—о, — момент размалывания пищи. Под кимо-граммой — отметка времени — 1 с.
Процесс жевания имеет четыре фазы — введения пищи в рот, ориентировочную, основную и формирования пищевого комка (рис. 11.10).
Регуляция жевания осуществляется рефлекторно. Возбуждение от рецепторов слизистой оболочки рта (механо-, хемо- и терморецепторов) передается по афферентным волокнам II, III ветви тройничного, языкоглоточного, верхнего гортанного нерва и барабанной струны в центр жевания, который находится в продолговатом мозге. Возбуждение от центра к жевательным мышцам передается по эфферентным волокнам тройничного, лицевого и подъязычного нервов. Возбуждение от чувствительных ядер ствола мозга по афферентному пути через специфические ядра таламуса переключается на корковый отдел вкусовой сенсорной системы, где осуществляется анализ и синтез информации, поступающей от рецепторов слизистой оболочки ротовой полости.
На уровне коры больших полушарий происходит переключение сенсорных импульсов на эфферентные нейроны, которые по нисходящим путям посылают регулирующие влияния к центру жевания продолговатого мозга.
Рефлекс слюноотделения.
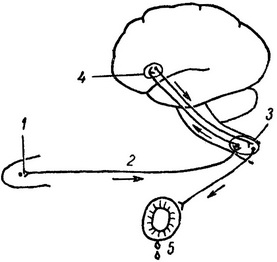 Рис.
52. Схема безусловного слюноотделительного
рефлекса.
Рис.
52. Схема безусловного слюноотделительного
рефлекса.
1 — вкусовой рецептор языка; 2 — чувствительный нерв; 3 — пищевой центр продолговатого мозга; 4 — пищевой центр коры; 5 — слюнная железа.
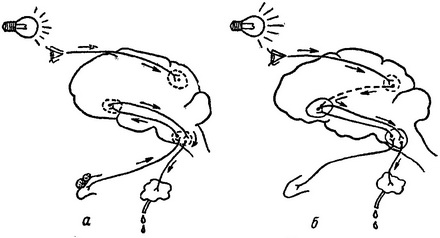 Рис.
53. Схема образования условного
слюноотделительного рефлекса.
Рис.
53. Схема образования условного
слюноотделительного рефлекса.
а — образование двух самостоятельных очагов возбуждения в коре: б — образование временной связи между двумя очагами возбуждения.
В качестве примера приведем один из опытов, поставленных в павловской лаборатории. Если перед кормлением собаки зажигать свет и многократно сочетать с ним дачу пищи, то через некоторое время слюна начинает выделяться у животного только при одном зажигании света. Чем это можно объяснить? При действии света происходит возбуждение зрительного центра коры больших полушарий, а под действием химических веществ пищи при кормлении возбуждается пищевой центр коры. При одновременном возбуждении двух участков коры больших полушарий между ними устанавливается связь. Для того чтобы такая связь установилась, нужно несколько сочетаний кормления с действием света. Когда эта связь возникла, возбуждение зрительного центра коры, воспринимающего световые раздражения, передается в слюноотделительный центр, вызывает его возбуждение и, таким образом, уже одно действие света вызывает выделение слюны. Индифферентный раздражитель (в нашем примере — свет) сигнализирует о предстоящем действии специфического раздражителя, который приводит в действие механизм безусловного рефлекса, поэтому система условных рефлексов рассматривается как сигнальная система — первая сигнальная система по И. П. Павлову. Она свойственна и животным, и человеку. Выделение слюны в ответ на световое раздражение может возникать только при определенных условиях, например при сочетании безусловного пищевого раздражителя с действием света, поэтому такой рефлекс и назван условным рефлексом. Аналогичным образом действует бесчисленное множество других индифферентных (неспецифических) раздражителей, в частности цвет, запах, общий вид пищи. Значит, выделение слюны при виде пищи — условный рефлекс. Так изучение механизма слюноотделения привело И. П. Павлова к открытию важнейших законов высшей нервной деятельности. Рассмотрим наиболее существенные черты условных рефлексов. Во-первых, условный рефлекс образуется только на основе врожденных рефлексов, т. е. только при сочетании действия раздражителя, вызывающего безусловный рефлекс, с любым другим. Например, можно получить выделение слюны при сочетании дачи пищи с прикосновением к коже, со звуком, с действием электрического тока, температурного фактора и т. п.
Позный тонус.
Одной из структур, ответственных за надсегментарный контроль позного тонуса, является красное ядро среднего мозга. Будучи составной частью организующей движения экстрапирамидной системы, красное ядро получает входы от моторной коры, ядер мозжечка, черной субстанции среднего мозга и дает начало руброспинальному тракту, который как общий путь обеспечивает регуляцию тонуса скелетных мышц.
Локальное электрическое раздражение крупноклеточной части красного ядра или самого руброспинального тракта вызывает возбуждение альфа- и гамма-мотонейронов мышц-сгибателей и одновременно торможение мотонейронов разгибателей. В этом отношении влияние красного ядра противоположно эффекту от раздражения вестибулоспинального тракта, который начинается от ядра Дейтерса. Вестибулоспинальный тракт оказывает возбуждающее действие на альфа- и гамма-мотонейроны разгибателей и тормозное — на мотонейроны сгибателей.
Статические и статокинетические рефлексы.
Рефлексы, вызываемые вестибулярными органами, можно разделить на две группы: так называемые статические и статокинетические рефлексы. Макулярные органы осуществляют статические рефлексы, которые поддерживают равновесие при разнообразных стоячих и наклонных положениях тела. Компенсаторное вращение глаз, особенно хорошо заметное у кошек, но возникающее и у людей, представляет собой статический рефлекс. Благодаря ему на сетчатке сохраняется изображение горизонтальных и вертикальных линий. Статокинетические рефлексы происходят во время движений. Один из них -это поворот, который происходит в свободном падении. Так, кошка всегда падает на лапы независимо от того, в каком положении она начала падать. Статокинетические рефлексы вызываются как макулярными органами, так и полукружными каналами. Другим статокинетическим рефлексом является "эффект лифта" на свободное падение, при котором усиливается тонус разгибателей, когда животное движется вниз. Среди статокинетических рефлексов особенно выделяется вестибулярный нистагм. Он состоит из последовательных движений глаз, вызываемых вестибулярной активностью, когда глаза движутся в сторону, противоположную вращению тела, благодаря чему направление взора остается неизменным. Это компенсаторное движение глаз, разумеется, эффективно только в определенных границах вращения. До того как глаза достигнут предела своего латерального движения, происходит их быстрое движение в сторону вращения -они устремляются вперед и фиксируются на новой точке. За этой быстрой фазой следует новое медленное движение, которое снова компенсирует вращение. Вращение головы или туловища вокруг вертикальной оси практически действует только на горизонтальные полукружные каналы. При этом отклонение купул в обоих горизонтальных каналах вызывает горизонтальный нистагм. Направление двух (быстрого и медленного) компонентов нистагма зависит от направления вращения (и, следовательно, от наклона купулы). При клиническом описании направление нистагма условились. Считать по быстрой фазе. Иными словами, при "правом нистагме" быстрая фаза направлена вправо. Такая терминология совпадает с терминами, принятыми для оптокинетического нистагма . При пассивном вращении первоначальный эффект состоит в стимуляции вестибулярного аппарата, а также относительном смещении видимого мира. Каждый из этих двух эффектов сам по себе вызывает нистагм (вестибулярный и оптокинетический). Они дополняют друг друга.
Ретикулярной формацией (РФ) называется сеть нейронов различных типов и размеров, имеющих многочисленные связи между собой, а также со всеми структурами ЦНС. Она располагается в толще серого вещества продолговатого, среднего и промежуточного мозга и регулирует уровень функциональной активности (возбудимости) всех нервных центров этих отделов ЦНС. Таким же образом она влияет на КБП.
В ЦНС выделяют две подсистемы, выполняющие разные организующие функции: специфическую и неспецифическую. Первая обеспечивает восприятие, проведение, анализ и синтез сигналов специфической чувствительности. К ним относятся все ее виды, т.е. зрительная, слуховая, болевая и т.д.
Неспецифической подсистемой является РФ. Она оказывает генерализованное возбуждающее или тормозящее влияние на многие структуры мозга. Следовательно, она может регулировать уровень функциональной активности моторной, сенсорной, висцеральных систем и организма в целом. Когда нервные импульсы идут по специфическим проводящим путям, по коллатералям этих путей они поступают и к нейронам РФ. Это приводит к их диффузному возбуждению. От нейронов РФ возбуждение передается на кору, что сопровождается возбуждением нейронов всех ее зон и слоев. Благодаря этому восходящему активирующему влиянию РФ, повышается активность аналитико-синтетической деятельности, увеличивается скорость рефлексов, организм подготавливается к реакции на неожиданную ситуацию. Поэтому РФ участвует в организации оборонительного, полового, пищедобывательного поведения. С другой стороны, она может избирательно активировать или тормозить определенные системы мозга. В свою очередь кора больших полушарий, через нисходящие пути, может оказывать возбуждающее действие на РФ.
Нисходящие ретикулоспинальные пути идут от РФ к нейронам спинного мозга. Поэтому она может оказывать нисходящие возбуждающие и тормозящие влияния на его нейроны. Например, ее гипоталамические и мезэнцефальные отделы повышают активность a-мотонейронов спинного мозга. В результате этого растет тонус скелетных мышц, усиливаются двигательные рефлексы. Тормозящее влияние РФ на спинальные двигательные центры осуществляется через тормозные нейроны Реншоу. Это приводит к торможению спинальных рефлексов.
РФ контролирует передачу сенсорной информации через продолговатый, средний мозг, а также ядра таламуса. Она непосредственно участвует в регуляции бодрствования и сна, за счет синхронизирующих центров сна и бодрствования, находящихся в ней. На нейроны РФ оказывают влияние различные фармакологические вещества: амфетамины, кофеин, LSD-25, морфин (опыт Эдисона).
Функции ретикулярной формации ствола мозга.
Ретикулярная формация - совокупность нейронов отростки которых образуют своеобразную сеть в пределах центральной нервной системы.
Ретикулярная формация открыта Дейтерсом, изучалась В. Бехтеревым, обнаружена в стволе мозга и спинном мозге. Основную роль выполняет ретикулярная формация ствола мозга. Ретикулярная формация занимает центральную часть на уровне продолговатого мозга, варолиевого моста, среднего и промежуточного мозга. Нейроны ретикулярной формации - клетки разнообразной формы, они имеют длинные ветвящиеся аксоны и длинные неветвящиеся дендриты. Дендриты образуют синапсы на нервных клетках. Некоторые дендриты выходят за пределы ствола мозга и доходят до поясничного отдела спинного мозга - они образуют нисходящий ретикулоспинальный путь.
Ретикулярная формация имеет связи с различными отделами центральной нервной системы: в ретикулярную формацию поступают импульсы от различных афферентных нейронов. Они поступают по коллатералям других проводящих путей. Ретикулярная формация не имеет непосредственных контактов с афферентной системой; ретикулярная формация имеет 2-х сторонние связи с нейронами спинного мозга - в основном с мотонейронами; с образованиями ствола мозга (с промежуточным и средним мозгом); с мозжечком, с подкорковыми ядрами (базальными ганглиями), с корой больших полушарий.
В ретикулярной формации ствола мозга различают 2 отдела:
растральный - ретикулярная формация на уровне промежуточного мозга;
каудальный - ретикулярная формация продолговатого мозга, моста и среднего мозга.
Изучены 48 пар ядер ретикулярной формации.
Функции ретикулярной формации
Функции ретикулярной формации изучены в 40-е гг. XX века Мэгуном и Моруции. Они проводили опыты на кошках, помещая электроды в различные ядра ретикулярной формации.
Ретикулярная формация обладает нисходящим и восходящим влиянием.
Нисходящее влияние - на нейроны спинного мозга. Оно (влияние) может быть активирующим и тормозным.
Восходящее влияние - на нейроны коры головного мозга - тоже тормозное и активизирующее. За счет особенности своих нейронов ретикулярная формация способна изменять функциональное состояние нейронов центральной нервной системы.
Особенности нейронов ретикулярной формации:
постоянная спонтанная электрическая активность - обеспечивается гуморальным влиянием и влиянием вышележащих отделов центральной нервной системы. Эта активность не имеет рефлекторного происхождения;
явление конвергенции - к ретикулярной формации идут импульсы по коллатералям различных проводящих путей. Сходясь к телам одних и тех же нейронов импульсы теряют свою специфичность; импульсы, поступая к нейронам ретикулярной формации, изменяют ее функциональную активность - если нейроны обладают выраженной электрической активностью, то под влиянием афферентных импульсов электрическая активность уменьшается и наоборот, т. е. модулируется активность нейронов ретикулярной формации; у нейронов ретикулярной формации низкий порог раздражения и, как следствие, высокая возбудимость; у нейронов ретикулярной формации высокая чувствительность к действию гуморальных факторов: биологически активных веществ, гормонов (адреналина), избытку СО2, недостатку О2 и т. д.;
в состав ретикулярной формации входят нейроны с различными медиаторами: адренэргические, холин-, серотонин-, дофаминэргические.
Дыхательный центр.
Дыхательный центр, совокупность нескольких групп нервных клеток (нейронов), расположенных в разных отделах центральной нервной системы, преимущественно в ретикулярной формации продолговатого мозга. Постоянная координированная ритмическая активность этих нейронов обеспечивает возникновение дыхательных движений и их регуляцию в соответствии с возникающими в организме изменениями. Импульсы от дыхательного центра поступают в двигательные нейроны передних рогов шейного и грудного отделов спинного мозга, от которых возбуждение передаётся к дыхательной мускулатуре. Активность дыхательного центра регулируется гуморально, т. е. составом омывающей его крови и тканевой жидкости, и рефлекторно, в ответ на импульсы, поступающие от рецепторов в дыхательной, сердечно-сосудистой, двигательной и др. системах, а также от высших отделов центральной нервной системы. При прекращении поступления нервных импульсов Дыхательный центр сохраняет активность (так называемый автоматизм дыхательного центра), способную обеспечить дыхательные движения.
Сосудодвигательный центр.
Сосудодвигательный центр локализован в продолговатом мозге и состоит из двух отделов — прессорного и депрессорного. Считают, что депрессорный отдел вызывает расширение сосудов, понижая тонус прессорного отдела и снижая, таким образом, эффект сосудосуживающих нервов. Влияния, идущие от сосудосуживающего центра продолговатого мозга, приходят к нервным центрам симпатической части вегетативной нервной системы (расположенным в боковых рогах грудных сегментов спинного мозга), регулирующим тонус сосудов отдельных участков тела. Кроме сосудодвигательных центров продолговатого и спинного мозга, на состояние сосудов оказывают влияние нервные центры промежуточного мозга и полушарий большого мозга.
Средний мозг.
Строение: Снизу граничит с задним, сверху с промежуточным мозгом. Имеет ножки и крышу(2 пары бугров четверохолмий). Ножки-из белого вещества, крыша и средняя часть-серое вещ-во.
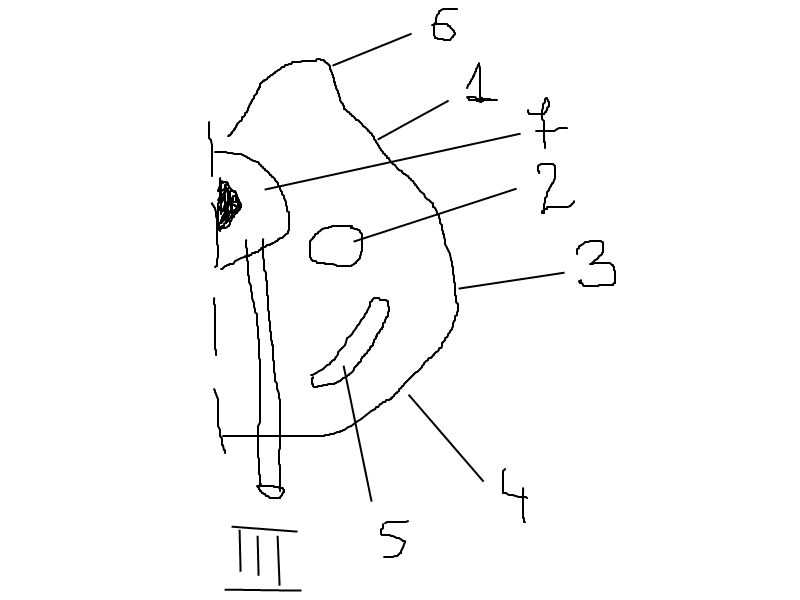
1.крыша
2.красное ядро
3.покрышка
4.ножки мозга
5.черное вещество
6.четверохолмия
7.центральное серое вещ-во
Рефлексы:сторожевой
1)Особое место среди этих рефлексов занимает так называемый четверохолмный рефлекс, анатомо-физиологическим субстратом которого является четверохолмие. Этот рефлекторный центр действует под влиянием всех рецепторов (зрительных, слуховых, проприоцептивных и т. д.). Свою деятельность он осуществляет главным образом через сетевидную формацию и регулируется условно рефлекторным влиянием коры головного мозга. Название «сторожевой» этот рефлекс получил потому, что он автоматически отвечает на всякое значительное внешнее раздражение, если оно наступает неожиданно. Внешне ответ выражается во вздрагивании, расширении зрачков, вскрикивании, быстром удалении раздражаемой части тела от источников раздражения, бегстве. Сторожевой рефлекс участвует во всей произвольной моторике. Угнетение этого рефлекса лишает человека возможности быстро включаться в движение.
Растормаживание сторожевого рефлекса происходит главным образом при нарушенной корковой нейродинамике и ослаблении тормозных влияний коры. В клинической практике рассматривают наблюдающуюся иногда общую эмоциональную реакцию (вздрагивание, вскрикивание), при вызывании сухожильных рефлексов, особенно коленных, - как признак растормаживания четверохолмного рефлекса.
2)ориентировочный рефлекс
ориентировочные рефлексы находятся на границе врожденной и приобретенной рефлекторной деятельности. С безусловными рефлексами их “роднит” то, что они носят врожденный характер, с условными – то, что они непостоянны и могут угасать. Ориентировочные рефлексы возникают на неожиданное действие каких-либо новых раздражителей окружающей среды и проявляются в ориентировочной реакции, получившей название “Что такое?”. В этом случае включается непроизвольное сенсорное внимание, что проявляется реакцией тревоги, настороженности и др. (при этом включается как вегетативные, так и соматические механизмы).
Более сложная форма этого рефлекса – ориентировочный рефлекс - имеет место на новую обстановку (ориентировочно-исследовательская деятельность). Любой вид аналитической деятельности происходит при активном участии этого рефлекса. В развитии ориентировочного рефлекса различают два этапа.
Первый этап проявляется в том, что прекращается текущая деятельность; второй этап проявляется увеличением двигательной активности, возникающей как следствие готовности организма к возможной встрече с раздражителем. Это проявляется увеличением тонуса мышц, возрастанием деятельности многих внутренних органов (дыхания, кровообращения и др.).
В развитии ориентировочного рефлекса различают два вида активности - локальную и генерализованную. Причем, в начале проявляется генерализованная, связанная в основном, с деятельностью ретикулярной формации ствола мозга, а затем - локальная, когда многие компоненты ориентировочного рефлекса постепенно угасают. Ориентировочным рефлексам отводится большое значение, так как они мобилизуют организм к неизвестной будущей деятельности.
основе образования любого условного рефлекса лежит универсальный механизм – механизм формирования временной связи между двумя возбужденными участками в ЦНС, которые возникают при действии условного и безусловного раздражителя, осуществляемый по типу проторения. Позже было показано, что в образовании временной связи большое значение имеет одновременная суммация и доминантное свойство, в результате которого более сильный очаг возбуждения в ЦНС, создаваемый безусловным раздражителем, притягивает к себе менее сильное раздражение, возникающее под влиянием условного сигнала.
3)Слуховой и слуховой рефлекс
Участие среднего мозга в регуляции движений и позного тонуса
Среднему мозгу принадлежит важная роль в регуляции глазных движений. Двигательный аппарат глаза состоит из шести наружных глазных мышц, которые иннервируются тремя черепными нервами .Нейроны ядра глазодвигательного нерва иннервируют медиальную, нижнюю и верхнюю прямые мышцы глаза, нижнюю косую, а также мышцу, поднимающую верхнее веко. Блоковый нерв иннервирует верхнюю косую мышцу, а отводящий нерв — латеральную прямую мышцу глаза. С помощью этого двигательного аппарата глаза могут производить горизонтальные, вертикальные и вращательные движения. При свободном рассматривании предметов, при чтении наши глаза совершают быстрые микросаккады (микроскачки), из одной точки фиксации в другую. Микросаккады чередуются периодами фиксации глаза, продолжающимися от 0,15 до 2 с. Горизонтальные движения глаза зависят от содружественной работы латеральной и медиальной прямых мышц глаза. степень возбуждения мотонейронов, локализованных в ядрах отводящего и глазодвигательного нервов, идущих к этим мышцам, контролируется центрами ретикулярной формации моста. В этих центрах обнаружены нейроны, которые характеризуются повышением частоты своей импульсной активности перед началом каждой горизонтальной микросаккады. Другая группа нейронов, напротив, прерывает свои импульсные разряды до и во время саккад.
Очевидно, характер разрядов нейронов ретикулярного центра определяет его активирующие или, наоборот, тормозные управляющие воздействия на мотонейроны отводящего и глазодвигательного нервов в соответствии с принципом антагонистической иннервации. Аналогичным образом при вертикальных движениях глаза также наблюдается антагонистическое взаимодействие между двумя другими группами глазных мышц. При движении глаза по вертикали вверх сокращаются нижняя косая и верхняя прямая мышцы (глазодвигательный нерв) и одновременно расслабляются верхняя косая (блоковый нерв) и нижняя прямая (глазодвигательный нерв) мышцы глаза.
Таким образом, ретикулярная формация среднего мозга играет важную роль в координации сокращений глазных мышц. Она получает афферентные входы от ростральных холмиков пластинки крыши, мозжечка, вестибулярных ядер, зрительных областей коры полушарий головного мозга. Поступающие по этим входам сигналы интегрируются центрами ретикулярной формации и служат для рефлекторного изменения работы глазодвигательного аппарата при внезапном появлении движущихся объектов, при изменении положения головы, при произвольных движениях глаз и т. д. По отношению к моторным центрам в ядрах черепных нервов ретикулярная формация выступает как более высокий уровень регуляции глазных движений, осуществляемой за счет возбуждающих и тормозных влияний.
Одной из структур, ответственных за надсегментарный контроль позного тонуса, является красное ядро среднего мозга. Будучи составной частью организующей движения экстрапирамидной системы, красное ядро получает входы от моторной коры, ядер мозжечка, черного вещества среднего мозга и дает начало красноядерно—спинномозговому пути, который как общий путь обеспечивает регуляцию тонуса скелетных мышц.
влияние красного ядра противоположно эффекту от раздражения преддверно—спинномозгового пути, который начинается от латерального преддверного ядра (ЛПЯ). Преддверно—спинномозговой путь оказывает возбуждающее действие на α— и γ—мотонейроны разгибателей и тормозное — на мотонейроны сгибателей.
Существует точка зрения, что красное ядро и ЛПЯ оказывают друг на друга тормозное воздействие, которое в норме снижает тонус антигравитационных разгибательных мышц. Если перерезку произвести на уровне переднего края ростральных холмиков и таким образом отделить красное ядро от заднего мозга, где локализовано ЛПЯ, то у кошки развивается децеребрационная ригидность.
Это состояние характеризуется резким повышением тонуса мышц—разгибателей конечностей, спины и хвоста (опистотонус). Оперированное животное, поставленное на лапы, сохраняет стоячее положение, так как сгибания в суставах не происходит. Если плоскость перерезки сдвинуть вперед и сохранить связь красного ядра с задним мозгом, то состояния децеребрационной ригидности не возникает. Децеребрационная ригидность устраняется при коагуляции ЛПЯ или при повторной перерезке стволовой части мозга каудальнее зоны расположения вестибулярных ядер.Все эти факты указывают на то, что основной причиной децеребрационной ригидности является снятие при перерезке тормозных влияний со стороны красного ядра на ЛПЯ. При этом начинает преобладать действие ЛПЯ, которое, как известно, возбуждает мотонейроны мышц—разгибателей.
Децеребрационная ригидность— рефлекторное состояние, оно поддерживается сенсорными сигналами от проприоцепторов мышц. Если перерезать дорсальные (чувствительные) корешки спинного мозга, то спазматическое сокращение разгибательных мышц исчезает. Очевидно, в естественных условиях красное ядро регулирует протекание проприоцептивных рефлексов спинного мозга, подчиняя их рефлексам более высокого порядка. Одним из путей этой регуляции является изменение возбудимости γ—мотонейронов, контролирующих чувствительность проприоцепторов мышц. Другой путь состоит в непосредственном воздействии на α—мотонейроны мышц—разгибателей.
Физиология мозжечка:
Структурная организация и связи мозжечка
У млекопитающих мозжечок — крупный вырост моста, состоящий из двух полушарий и непарного отдела — червя. Со стволовой частью мозга мозжечок соединяется тремя парами ножек. Самые толстые средние ножки как бы охватывают продолговатый мозг и, расширяясь, переходят в мост. Ростральные ножки начинаются в зубчатых ядрах мозжечка и направляются к пластинке крыши среднего мозга. Третья пара ножек (каудальная) спускается вниз, сливаясь с продолговатым мозгом. Афферентные волокна, приходящие в мозжечок, преимущественно входят в состав средних и каудальных ножек, тогда как эфферентные собраны главным образом в ростральных ножках мозжечка.
Вся поверхность мозжечка разделяется глубокими бороздами на доли. В свою очередь, каждая доля параллельными бороздками разделяется на извилины;группы извилин формируют дольки мозжечка.
Наиболее изолированная клочково—узелковая доля (X) составляет древнюю часть мозжечка (палеоцеребеллум). Здесь заканчиваются проекции от вестибулярных ядер продолговатого мозга. Следующий отдел — старая часть мозжечка, или археоцеребеллум, — включает в себя участки червя, соответствующие ростральной доле, а также пирамиду, язычок и околоклочок. В палеоцеребеллуме находятся проекции восходящих спинно—мозжечковых путей, несущих информацию от мышечных рецепторов. И наконец, третий отдел — новая часть мозжечка, или неоцеребеллум, — состоит из появляющихся у млекопитающих полушарий и участков червя, которые расположены каудальнее первой щели. К неоцеребеллуму по путям, переключающимся в ядрах моста, поступает афферентная импульсация от обширных областей коры больших полушарий (лобных, теменных, височных и затылочных долей).
Полушария и червь мозжечка состоят из лежащего на периферии серого вещества — коры. — и расположенного глубже белого вещества — мозгового тела, в котором заложены скопления нервных клеток, образующие ядра мозжечка. Кора мозжечка представлена тремя слоями .Самый поверхностный слой — молекулярный — состоит из параллельных волокон и разветвлений дендритов и аксонов нейронов нижележащих слоев. В нижней части молекулярного слоя расположены тела корзинчатых нейронов, аксоны которых оплетают тела и начальные сегменты аксонов клеток Пуркинье (грушевидных нейронов) . Здесь же в молекулярном слое имеется некоторое количество звездчатых клеток.
Вентральное молекулярного слоя находится слой грушевидных нейронов, в котором сосредоточены тела клеток Пуркинье .Их дендриты поднимаются вверх и широко ветвятся в молекулярном слое. Дендриты клеток Пуркинье содержат множество шипиков, на которых образуют синапсы параллельные волокна молекулярного слоя. Аксоны клеток Пуркинье спускаются к ядрам мозжечка. Часть из них заканчивается на вестибулярных ядрах. Практически аксоны клеток Пуркинье представляют собой единственный выход из коры мозжечка.
Под ганглиозным слоем лежит зернистый слой, который содержит большое число зерен, или гранулярных клеток. Аксоны клеток—зерен поднимаются вертикально вверх, в молекулярный слой и там Т—образно ветвятся. Ветви идут параллельно поверхности коры и образуют синапсы на дендритах других клеток.
Афферентный вход к нейронному аппарату коры осуществляется по трем системам нервных волокон. Это, во—первых, лазающие, или ползучие, волокна, идущие из нижних олив продолговатого мозга. Нижняя олива получает афференты от нескольких восходящих путей спинного мозга и из центров головного мозга. Лазающие волокна широко ветвятся и подобно лианам оплетают дендриты клеток Пуркинье, формируя на них синапсы.
Вторая система афферентных волокон — это моховидные, или мшистые, волокна, идущие от ядер моста и оканчивающиеся на клетках—зернах. Мшистые волокна многократно ветвятся и образуют синапсы на множестве клеток коры мозжечка. И наконец, третья система афферентных волокон — это также широко ветвящиеся адренергические волокна, поступающие в кору мозжечка из голубоватого пятна в среднем мозгу. Оно представляет собой скопление из нескольких нейронов, аксоны которых способны диффузно выбрасывать норадреналин в межклеточное пространство. Вероятно, эти нейроны выполняют нейромодуляторную функцию и могут изменять возбудимость нейронов, локализованных в коре мозжечка.
Таким образом, большинство связей, опосредованных интернейронами коры мозжечка, являются тормозными. Исключение составляют только клетки—зерна, которые возбуждаются от моховидных волокон и сами через Т—образно ветвящиеся аксоны активируют все остальные интернейроны коры мозжечка. Однако конечный эффект этой активации опять—таки сводится к торможению.