

XII.2.5.4. Производные гетероциклов с двумя и более гетероатомами. Нуклеиновые кислоты.
Среди подобных соединений наиболее важны производные пиримидина и конденсированных систем, включающих пиримидиновый цикл, прежде всего, пурина.
Производными пурина являются известные алкалоиды кофеин (923), содержащийся в больших количествах в листьях чая и теобромин (924), содержащийся в бобах какао. Известный природный фактор роста – фолиевая кислота (925) – производное птеридина. Производными конденсированной трициклической системы –бензо[g]птеридина –являются рибофлавин (витамин В2) (926) и важный кофермент флавинмононуклеотид (FMN) (927):
 Наиболее
важные природные соединения, включающие
гетероциклические системы с двумя и
более атомами азота, – нуклеиновые
кислоты (НК).
Их основные биологические функции –
хранение, копирование и передача
генетической информации.
Наиболее
важные природные соединения, включающие
гетероциклические системы с двумя и
более атомами азота, – нуклеиновые
кислоты (НК).
Их основные биологические функции –
хранение, копирование и передача
генетической информации.
Нуклеиновые кислоты – линейные биополимеры; их цепь построена из чередующихся пентозных и фосфатных фрагментов, связанных по типу полиэфира; боковыми группами являются производные пиримидина и пурина – их часто называют гетероциклическими основаниями (на схеме обозначены как B – Base):
 В
состав нуклеиновых кислот входят
фрагменты двух альдопентоз – D-рибозы
(Х=ОН) и D-2-дезоксирибозы
(Х=Н); первой – в состав рибонуклеиновых
кислот (РНК),
второй – в состав дезоксирибонуклеиновых
кислот (ДНК).
Пентозы входят в структуры НК в фуранозной
форме.
В
состав нуклеиновых кислот входят
фрагменты двух альдопентоз – D-рибозы
(Х=ОН) и D-2-дезоксирибозы
(Х=Н); первой – в состав рибонуклеиновых
кислот (РНК),
второй – в состав дезоксирибонуклеиновых
кислот (ДНК).
Пентозы входят в структуры НК в фуранозной
форме.
В качестве боковых групп (гетероциклических оснований) в состав нуклеиновых кислот входят фрагменты пяти соединений: урацила (В1), тимина (В2), цитозина (В3), аденина (В4) и гуанина (В5); первые три – производные пиримидина, последние два – производные пурина. Связь между пентозным и гетероциклическим фрагментами осуществляется по типу N-гликозида; в образовании этой связи участвуют атом N1 пиримидиновых оснований и атом N9 пуриновых оснований. В состав любой нуклеиновой кислоты входят только четыре из пяти гетероциклических оснований: в состав ДНК не входит урацил, в состав РНК – тимин.
Фосфатные группы связывают фрагменты пентоз по типу 3’,5’-фосфодиэфирных связей. В физиологических условиях фосфатные гидроксигруппы ионизированы (Р-ОН Р-О¯ ).
На одном конце полимерной цепи находится звено, содержащее фосфатную группу, соединенную с положением 5’ пентозы; это концевое звено называют 5’-концом; на другом конце – звено, содержащее свободную 3’-ОН – группу; это звено называют 3’-концом. [Положения пентозы обозначают цифрами со штрихом; цифрами без штриха обозначают положения в гетероциклических основаниях].
Элементарные звенья нуклеиновых кислот называют нуклеотидными звеньями; им соответствуют мономеры – нуклеотиды. Нуклеотид включает фрагмент пентозы, гетероциклическое основание и остаток фосфорной кислоты; ниже приведены два примера:
В природных нуклеотидах фосфорилировано
положение 5’ пентозы (а не положение
3’); такие нуклеотиды называют
нуклеозид-5’-фосфатами
(нуклеозид
– N-гликозид,
образованный из пентозы и гетероциклического
основания, см. стр. 288). Если нуклеотид
включает D-2-дезоксирибозу
[как в соединении (928)], то он называется
дезоксирибонуклеотидом,
если D-рибозу
[как в соединении (929)], то рибонуклеотидом.
природных нуклеотидах фосфорилировано
положение 5’ пентозы (а не положение
3’); такие нуклеотиды называют
нуклеозид-5’-фосфатами
(нуклеозид
– N-гликозид,
образованный из пентозы и гетероциклического
основания, см. стр. 288). Если нуклеотид
включает D-2-дезоксирибозу
[как в соединении (928)], то он называется
дезоксирибонуклеотидом,
если D-рибозу
[как в соединении (929)], то рибонуклеотидом.
 Полинуклеотидная
цепь может образовываться путем
поликонденсации нуклеотидов: группа
С3’-ОН
предыдущего
звена образует эфирную связь с
5’-фосфатной группой последующего
звена ( если вести синтез с 5’-конца, как
это и происходит в природе):
Полинуклеотидная
цепь может образовываться путем
поликонденсации нуклеотидов: группа
С3’-ОН
предыдущего
звена образует эфирную связь с
5’-фосфатной группой последующего
звена ( если вести синтез с 5’-конца, как
это и происходит в природе):
Естественно, таким путем могут образовываться и олигонуклеотиды: динуклеотиды (два нуклеотидных звена), тринуклеотиды (три звена) и т.д.
 Приведенная
схема имеет скорее формальный характер:
при лабораторных
синтезах олиго- и полинуклеотидов
используют модифицированные нуклеозиды
и нуклеотиды, а при биосинтезе
нуклеиновых кислот мономерами являются
не нуклеотиды, а нуклеозид-
5’-трифосфаты –
аналоги нуклеотидов, содержащие фрагменты
трифосфорной кислоты; в качестве примера
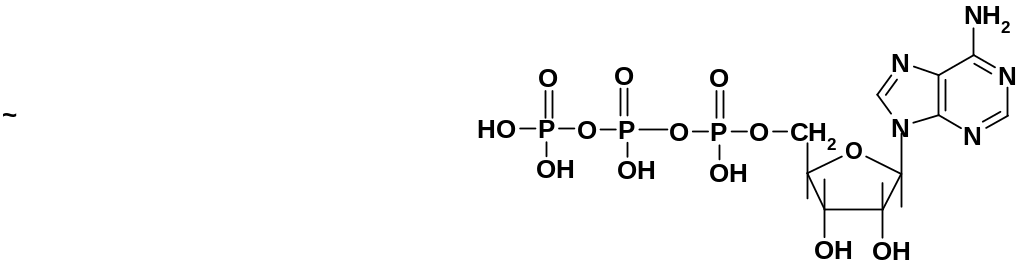
можно привести аденозин-5’-трифосфорную
кислоту (АТФ):
Приведенная
схема имеет скорее формальный характер:
при лабораторных
синтезах олиго- и полинуклеотидов
используют модифицированные нуклеозиды
и нуклеотиды, а при биосинтезе
нуклеиновых кислот мономерами являются
не нуклеотиды, а нуклеозид-
5’-трифосфаты –
аналоги нуклеотидов, содержащие фрагменты
трифосфорной кислоты; в качестве примера
можно привести аденозин-5’-трифосфорную
кислоту (АТФ):
Синтез полинуклеотидной цепи из этих мономеров идет по той же схеме, что и из нуклеотидов, но в ходе процесса выделяется не вода, а пирофосфорная кислота (точнее – пирофосфат). Нуклеозидтрифосфаты более активны, чем нуклеотиды, т.к. связи Р-О-Р имеют высокую энергию и очень легко подвергаются алкоголизу в условиях синтеза.
Молекулярная масса нуклеиновых кислот варьирует в очень большом диапазоне. Самые низкомолекулярные – транспортные РНК – имеют молекулярную массу порядка 3·104; самые высокомолекулярные – хромосомные ДНК эукариот – до 1010.
Нуклеиновые кислоты, как и белки, являются монодисперсными сополимерами, обладающими первичной структурой. Как и в белках, полимерная цепь является стандартной и монотонно построенной; специфика звеньев определяется боковыми группами – гетероциклическими основаниями. Существование специфической первичной структуры позволяет накапливать огромный объём информации, несмотря на то, что первичная структура образована всего четырьмя типами звеньев («четырехбуквенный алфавит», в отличие от белкового –«двадцатибуквенного»).
Как и белки, нуклеиновые кислоты – полимеры с жесткими макромолекулами; для них характерны весьма сильные внутри- и межмолекулярные взаимодействия, из которых особо следует отметить водородные связи между фрагментами гетероциклических оснований. В физиологических условиях нуклеиновые кислоты конформационно стабильны; как и белки, они имеют не только первичную, но и вторичную и третичную структуры.
Дезоксирибонуклеиновые кислоты (ДНК) – хранители генетической информации, закодированной в первичных структурах ДНК. Вторичная структура большинства ДНК – двойная спираль, образованная фрагментами двух цепей, ориентированных антипараллельно (одна в направлении от 5’-конца к 3’-концу, другая – в обратном направлении) (рис. 10)
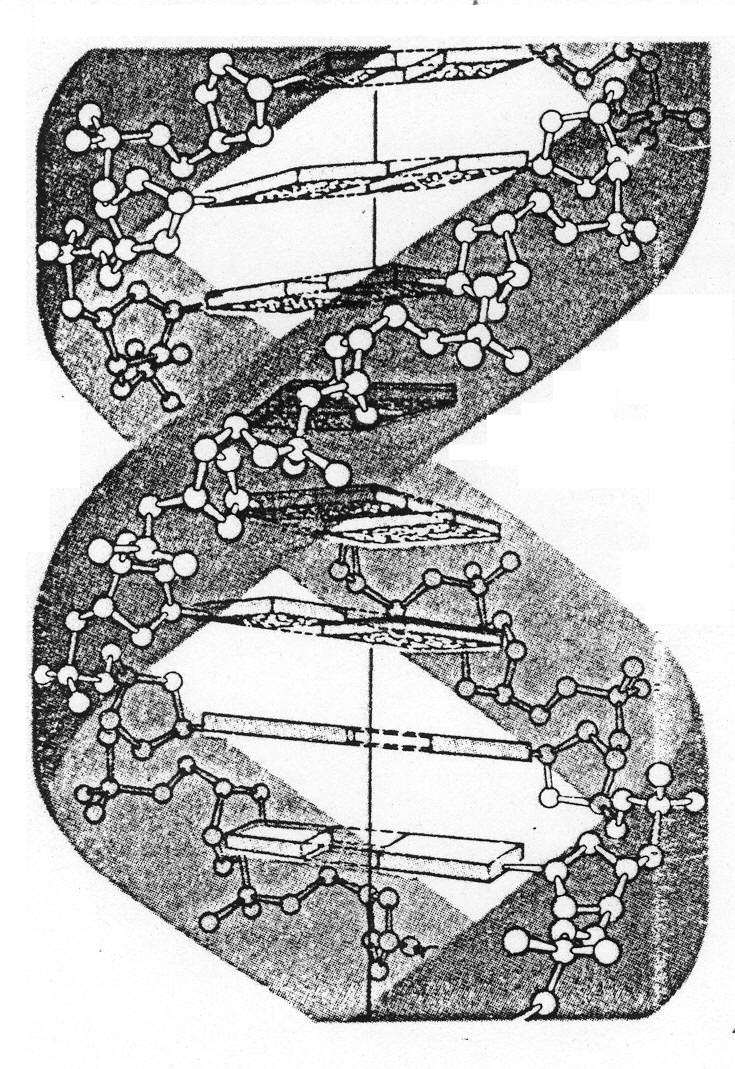
Рис. 10. Схема построения двойной спирали ДНК.
Структура двойной спирали энергетически весьма выгодна; главную роль при этом играют два вида взаимодействий:
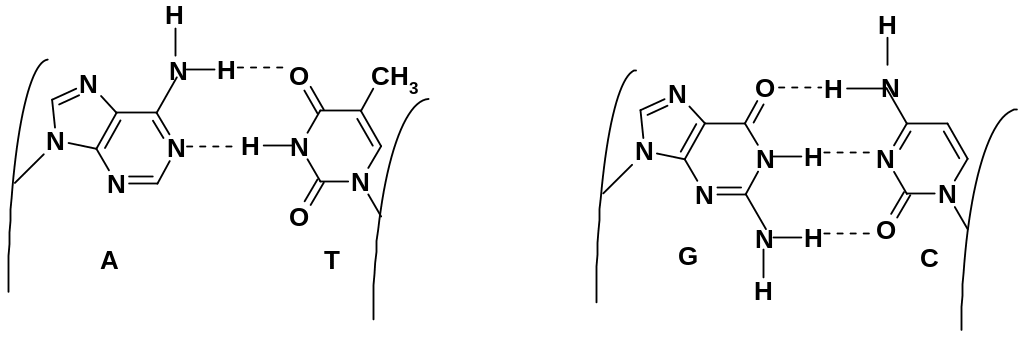 1.
Межцепные
водородные связи
между фрагментами гетероциклических
оснований; при этом образуются связи
только в двух парах оснований: аденин-тимин
(A-T)
и гуанин-цитозин (G-C).
В первой паре образуются две водородных
связи, во второй –три:
1.
Межцепные
водородные связи
между фрагментами гетероциклических
оснований; при этом образуются связи
только в двух парах оснований: аденин-тимин
(A-T)
и гуанин-цитозин (G-C).
В первой паре образуются две водородных
связи, во второй –три:
Таутомерные формы оснований обеспечивают их прочное связывание водородными связями. Основания в этих парах столь точно соответствуют друг другу, что длины обеих межцепных «мостиков», образованных парами оснований, абсолютно одинаковы (1,085 нм). Точное структурное соответствие – комплементарность в парах аденин-тимин и гуанин-цитозин обеспечивает не только построение двойной спирали, но и возможность матричного синтеза на макромолекулах ДНК.
2. Взаимодействие между параллельно расположенными парами оснований. В двойной спирали пары плоских оснований расположены параллельно друг над другом на достаточно близком расстоянии («сложены стопкой»). Между параллельными парами оснований возникают силы Ван-дер-Ваальса, которые дополнительно стабилизируют структуру: эти взаимодействия называют стэкинг-взаимодействиями. Стэкинг-взаимодействия стабилизируют двойную спираль «по вертикали», в то время, как водородные связи – «по горизонтали».
Рибонуклеиновые кислоты (РНК) осуществляют передачу генетической информации от ДНК к белкам. Известны три типа РНК: 1. Матричные (информационные), (мРНК); они непосредственно воспринимают информацию от ДНК и передают ее на белки посредством генетического кода; 2. Рибосомные (рибосомальные) (рРНК); они вместе с белками образуют внутриклеточные органеллы – рибосомы, на которых происходит трансляция – матричный синтез белков на матричной РНК; 3. Транспортные (тРНК); они обеспечивают связывание аминокислот с матричной РНК при трансляции.
В отличие от большинства ДНК, которые являются двухцепочечными, РНК существуют в виде одиночных цепей. Вторичная структура наиболее изучена для транспортных РНК; она представляет собой «клеверный лист» (рис. 11). Эта структура стабилизируется внутрицепными водородными связями в парах комплементарных оснований аденин-урацил (А-U) и гуанин-цитозин (G-C).
Вторичные структуры рибосомных РНК намного более сложны, но вероятнее всего построены по тем же принципам, что и для транспортных РНК.
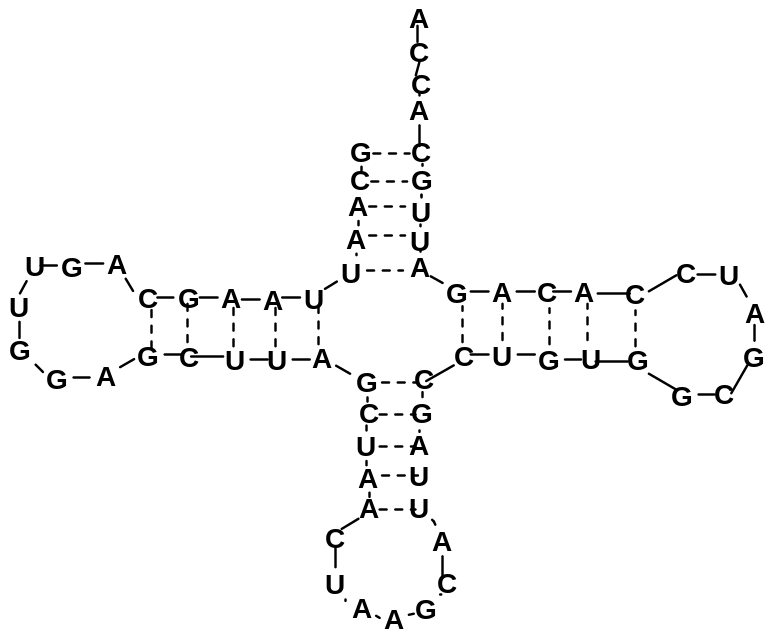 Рис.11. «Клеверный
лист» аланиновой тРНК из дрожжей.
Рис.11. «Клеверный
лист» аланиновой тРНК из дрожжей.
Уникальность структуры нуклеиновых кислот не только в том, что они могут содержать огромное количество химической информации, но, прежде всего, в том, что они способны размножать (тиражировать) эту информацию и передавать ее на другие химические объекты – белки. Эти едва ли не самые важные биологические функции осуществляются путем матричного синтеза: макромолекула нуклеиновой кислоты служит матрицей для синтеза другой полимерной цепи, передавая ей свою информацию. Возможность такого считывания информации в ходе синтеза базируется на комплементарности пар гетероциклических оснований в нуклеиновых кислотах: в ходе синтеза гетероциклические основания мономеров «узнают» комплементарные им основания на молекуле-матрице; синтезированная цепь комплементарна матрице (в виде упрощенной схемы показан принцип синтеза макромолекулы ДНК из мономеров в процессе репликации):
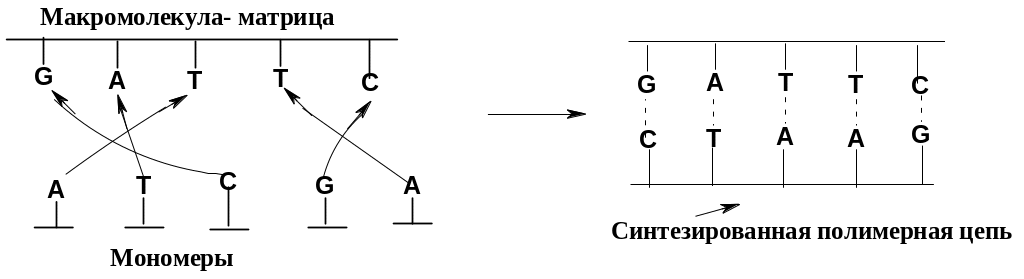 Матричный
синтез (наряду с ферментативным катализом)
– наиболее совершенное воплощение
принципа молекулярного
распознавания,
простейшие варианты которого
рассматривались на примере краун-эфиров.
Матричный
синтез (наряду с ферментативным катализом)
– наиболее совершенное воплощение
принципа молекулярного
распознавания,
простейшие варианты которого
рассматривались на примере краун-эфиров.
Матричный синтез полимеров осуществляется в ходе трех важнейших генетических процессов: репликации, транскрипции и трансляции.
Репликация – процесс копирования генетической информации. В процессе репликации двойная спираль ДНК расплетается, освобождая единичные цепи, каждая из которых служит матрицей для синтеза из мономеров (дезоксирибонуклеозидтрифосфатов) комплементарной ей цепи ДНК. В результате из одной двухцепочечной макромолекулы образуются две идентичных; количество ДНК удваивается.
Транскрипция – процесс передачи информации от ДНК к РНК; здесь также происходит расплетение двойной спирали ДНК, но только одна одиночная цепь служит матрицей, причем для синтеза не цепи ДНК, а цепи РНК ( здесь мономерами служат рибонуклеозидтрифосфаты).
Трансляция – процесс передачи информации на белок. Здесь мРНК служит матрицей синтеза уже не полинуклеотидной, а полипептидной цепи. Для прохождения этого синтеза нужен посредник (адаптор); таким посредником является тРНК: она узнаёт, с одной стороны, «свою» аминокислоту, а с другой - тот участок мРНК, который кодирует эту аминокислоту (каждая аминокислота кодируется кодоном – специфическим участком мРНК, содержащим три звена).
Репликация, транскрипция и трансляция являются сложнейшими биохимическими процессами, включающими множество стадий и катализируемыми целыми комплексами ферментов. Даже не слишком подробное описание этих процессов выходит за рамки данного курса. Но ключевые моменты всех этих процессов – образование пар комплементарных оснований, как было показано выше.
Матричный синтез – один из наиболее совершенных видов органического синтеза: это программируемый синтез, позволяющий конструировать желаемую молекулу максимально целенаправленно; особо важен матричный синтез для получения полимеров с заданными свойствами. Матричные синтезы в живом организме пока остаются недосягаемым образцом для химиков; тем не менее, определенные успехи достигнуты. Одним из них является метод «репликации в пробирке» – полимеразная цепная реакция (К.Маллис, 1987); этот метод позволяет многократно копировать большие фрагменты ДНК вне организма (используя мономерные нуклеозидтрифосфаты, фермент ДНК-полимеразу и олигонуклеотидные «затравки» – праймеры). Метод весьма прост по исполнению.
Резюме
Шестичленные ароматические гетероциклы, не имеющие заряда, включают атомы азота – один или более. Шестичленная гетероароматическая система, содержащая атом кислорода, может быть только катионной.
Шестичленные гетероароматические соединения являются дефицит- ными; они реагируют с электрофильными реагентами труднее бензоидных ароматических соединений, а с нуклеофильными реагентами - легче. Дефицитность растет при переходе от пиридина к диазинам и особенно сильно проявляется в солях пирилия.
Пиридин и диазины могут алкилироваться по атому азота, образуя четвертичные соли; при действии надкислот они образуют N-оксиды, которые заметно легче реагируют с электрофильными реагентами, чем исходные гетероциклы. Пиридин проявляет свойства довольно слабого основания; еще менее основны диазины.
Шестичленные гетероциклы повышают СН-кислотность связанных с ними метильных и метиленовых групп, которые вследствие этого могут играть роль метиленовых компонент в реакциях конденсации с карбонильными соединениями. Для многих гидроксипроизводных пиридина и диазинов наблюдается переход в таутомерную лактамную форму; для аминопроизводных переход в иминную форму не наблюдается.
Бензоконденсированные производные пиридина и диазинов во многом аналогичны по свойствам соответствующим одноядерным гетероциклам, но в ряде случаев электрофильное замещение идет не по гетероциклическому, а по бензольному ядру.
Производные как одноядерных, так и конденсированных гетероцикли- ческих систем широко распространены в живой природе как алкалоиды, коферменты, факторы роста. Наиболее важными природными соединениями, включающими гетероциклические структуры, являются нуклеиновые кислоты. Решающую роль в их биологических функциях играют гетероциклические фрагменты: они придают устойчивость нативным конформациям нуклеиновых кислот и обеспечивают комплементарность звеньев; это позволяет осуществлять матричный синтез и таким путем копировать и передавать наследственную информацию.