

6. Гормоны поджелудочной железы
Поджелудочная железа состоит из двух функционально различных частей: экзокринной и эндокринной. В экзокринной части, которая составляет 97-95% массы железы, вырабатываются пищеварительные ферменты (рис. 6.1).
Рис. 6.1. Положение поджелудочной железы.
Э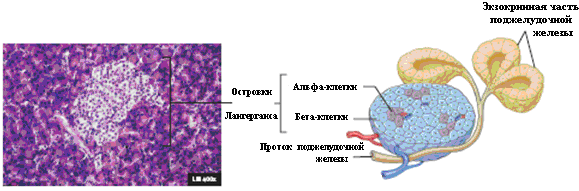 ндокринная
часть представлена островками клеток,
которые вкраплены в экзокринную часть.
По имени автора, описавшего эти островки
в 1869 г., они получили название островков
Лангерганса (рис. 6.2). В поджелудочной
железе сотни тысяч островков, но они
составляют всего лишь 2-3% общей массы
железы.
ндокринная
часть представлена островками клеток,
которые вкраплены в экзокринную часть.
По имени автора, описавшего эти островки
в 1869 г., они получили название островков
Лангерганса (рис. 6.2). В поджелудочной
железе сотни тысяч островков, но они
составляют всего лишь 2-3% общей массы
железы.
Рис.6.2. Гистология поджелудочной железы.
Островки Лангерганса состоят из четырех видов клеток, которые вырабатывают глюкагон (А (альфа)-клетки), инсулин (В (бета)-клетки, соматостатин (D (дельта)-клетки) и панкреатический полипептид (РР) (F-клетки) (табл.6.1).
Таблица 6.1.
Типы клеток островков Лангерганса и синтезируемые в них гормоны
-
Тип клеток
Гормоны
Процентное содержание
А-клетки (альфа)
В-клетки (бета)
D-клетки (дельта)
F-клетки
Глюкагон
Инсулин
Соматостатин
Панкреатический полипептид
По периферии островков 20-25
75-80
Располагаются в их центральной части
По периферии островков <1
По периферии островков <1
История открытия инсулина
Известный английский врач Т. Виллис (который, кстати, был и одним из учредителей Лондонского королевского общества) славился своей любознательностью. В стремлении выяснить истину его ничто не могло остановить. Именно он впервые связал развитие диабета с повышенным уровнем сахара в организме. Прибором для этого ему послужил один из самых надежных и чувствительных аппаратов – собственный язык. Попробовав на вкус мочу диабетиков, Виллис убедился в том, что она сладкая. Но на эту находку Виллиса как-то не обратили должного внимания, восприняв ее как причуду почтенного медика. И только через 100 лет после Виллиса другой английский врач П. Добсон установил, что в моче диабетических больных содержится сахар – глюкоза.
Немецкие ученые И. Меринг и О. Минковски занимались изучением роли поджелудочной железы в процессе пищеварения. Каково же было их удивление, когда однажды утром, придя на работу и заглянув в операционную, где с вечера была оставлена собака, у которой накануне удалили поджелудочную железу, экспериментаторы увидели, что она вся была облеплена мухами. Осмотрев животное, они поняли, что мух привлекал сахар, в избытке содержащийся в моче собаки. Предприняв, теперь уже специальные, исследования, немецкие ученые в 1889 году убедительно показали, что у собак с удаленными поджелудочными железами развиваются все признаки сахарного диабета, приводящие их к скорой смерти.
Первые попытки выделить неизвестный гормон из островков Лангерганса успеха не имели. Однако эти неудачи тоже внесли свой вклад в будущие открытия. Именно благодаря им ученые предположили, что гормон должен иметь белковую (пептидную) природу, поскольку причиной неудачного выделения можно было считать возможность разрушения искомого белка собственными протеолитическими ферментами, поджелудочной железы.
В 1900 году всю проблему уже можно было бы разрешить. Тогда русский исследователь Соболев проделал хорошо продуманный опыт. Поджелудочная железа выделяет через выводной проток в тонкую кишку сок, столь важный для пищеварения. Соболев перевязал у собаки этот проток, после чего железистая ткань, которая стала излишней, сморщилась. Несмотря на это, животное не заболело диабетом. Очевидно, заключил ученый, в железе что-то сохранилось, и этот остаток предотвратил возникновение сахарной болезни. При вскрытии трупа животного он нашел в железе клетки Лангерганса. Они, как можно было заключить, и представляют собой орган, который регулирует количество сахара в организме. Открытие Соболева вначале оставалось неизвестным ученому миру, так как было описано лишь в русской литературе.
29-летний сотрудник университета Западного Онтарио (Канада) Фредерик Бантинг. Ф. Бантинг, повторивший опыт Соболева, убедился в том, что действительно при нарушении протока поджелудочной железы островки Лангерганса сохраняются. Тогда он решил попытаться выделить гормон не из нормальных, а из тех поджелудочных желез, у которых были перевязаны протоки, предохраняя тем самым инсулин от ферментативного расщепления.
Свою идею Бантинг рассказал известному канадскому естествоиспытателю Дж. Маклеоду – руководителю кафедры физиологии университета в Торонто. Маклеод горячо поддержал намерения Бантинга и вместе со своей хорошо оснащенной лабораторией предоставил в его распоряжение помощника – студента 5-го курса Чарлза Беста – хорошо зарекомендовавшего себя молодого исследователя, искусно владевшего химическими методами определения сахара в крови. Успех пришел быстро. Уже в августе 1921 года они получили очищенные препараты гормона и убедились в его сильном лечебном действии на собаке, страдавшей тяжелой формой экспериментального диабета. Вскоре исследователи научились выделять инсулин из поджелудочных желез телят и коров. Фармацевтические заводы стали производить в больших количествах этот гормон, получая сырье для него на мясокомбинатах. Сотни тысяч больных смогли пользоваться мощным средством борьбы с тяжким недугом.
В 1923 году за выдающиеся исследования Ф. Бантинг и Дж. Маклеод получили Нобелевскую премию. Парлоз Бест не был выдвинут на премию, а Маклеод не принил участие в исследованиях, тогда как другой его сотрудник Джон Колип разработал другой способ получения И. Бантинг и Маклеод публично заявили о сложившейся ситуации, но менять решениея Нобелевского комитета нельзя. Тогда свою часть премии Бантинг поделил с Бестом. Маклеод с Компом после открытия инсулина Фредерик Бантинг проработал только 18 лет... В самом расцвете творческих сил трагически оборвалась жизнь выдающегося ученого, которому миллионы людей обязаны жизнью. В 1941 году в возрасте 50 лет он погиб в авиационной катастрофе.
Первое введение инсулина больному сахарным диабетом было осуществлено в январе 1923 года – через 17 месяцев после открытия гормона. Такой короткий срок может служить примером, достойным подражания при внедрении современных результатов исследования в практику здравоохранения. С тех пор инсулин лечит больных. А больных, нуждающихся в нем, много. Недаром сахарный диабет называют «болезнью цивилизации».
Несмотря на сильный лечебный эффект инсулина, его применение весьма ограничено. Во-первых, потому, что действует он только при введении в кровь (то есть инъекциях), и при этом лечебный эффект сохраняется лишь в течение 4-6 часов. Затем инъекции необходимо повторять. Во-вторых, при передозировке инсулина возникает серьёзное осложнение – гипогликемия (резкое падение содержания сахара). А снизить дозу инсулина – значит не получить желаемого лечебного эффекта.
Без инсулина организм существовать не может. И дело не в том, что при этом происходит накопление сахара со всеми вытекающими отсюда трагическими последствиями, но и в необходимости инсулина для обеспечения самых разных физиологических процессов, начиная от регуляции обмена углеводов и кончая клеточным делением, развитием и размножением живых организмов.
Биосинтез инсулина
У человека ген инсулина локализуется в коротком плече 11-й хромосомы. Предшественником молекулы инсулина является препроинсулин, который синтезируется в эндоплазматическом ретикуломе В-клеток. Сразу же, под влиянием протеолитических энзимов, препроинсулин превращается в проинсулин и транспортируется в аппарат Гольджи, где запасается в виде гранул. Там же протеолитические энзимы расщепляют проинсулин в равных количествах на инсулин и С-пептид, с остатком небольшого количества проинсулина.
Проинсулин состоит из 86 аминокислот, в виде одной цепи с молекулярные весом (MB) 9000. В него входят А- и В-цепи инсулина и соединяющий их С-пептид. Проинсулин имеет слабо выраженную биологическую активность, секретируется в кровь в небольших количествах и составляет 3-5% иммунореактивного сулина в плазме крови. Так как период полураспада проинсулина в 3-4 раза больше, чем инсулина, он накапливается в плазме крови и, в конечном итоге, составляет 12-20% циркулирующего иммунореактивного инсулина.
С-пептид, соединяющий А- и В-цепи инсулина в проинсулине, состоит из 31 аминокислоты в виде одной цепи. Он не обладает биологической активностью, и освобождается в кровь в равных количествах с инсулином. В связи с этим по концентрации С-пептида в плазме крови можно судить о синтетической и секреторной активности В-клеток островков. Период полураспада С-пептида в 3-4 раза длиннее, чем инсулина, так как он не катаболизируется в печени. Поэтому в базальном состоянии (натощак) концентрация С-пептида в плазме крови значительно превышает концентрацию инсулина (в 10-15 раз) и может достигать 1000 пмоль/л (5-6 нг/мл).
Химическая структура инсулина расшифрована в 1958г. Сангером, за что был удостоен Нобелевской премии.
Инсулин состоит из 51 аминокислоты в виде двух цепей, соединенных двумя дисульфидными мостиками. Короткая цепь, или цепь А, состоит из 21 аминокислоты, и цепь В, или длинная цепь, – из 30 аминокислот. Один дисульфидный мостик расположен между седьмыми аминокислотами обеих цепей, другой соединяет 20-ю аминокислоту короткой цепи и 19-ю аминокислоту длинной цепи. Кроме того, в короткой цепи инсулина имеется дисульфидный мостик между 6-й и 11-й аминокислотами. Молекулярный вес человеческого инсулина 5808. По своему составу человеческий инсулин ближе всего к свиному инсулину и отличается от последнего лишь одной аминокислотой. У человека 30-я аминокислота длинной цепи – треонин, а в свином инсулине – аланин.
У взрослого человека секретируется в кровь 30-40 ЕД инсулина в сутки. Период полураспада инсулина составляет 2-3 мин. Катаболизм происходит главным образом в печени и в почках.
В течение многих десятилетий единственным источником инсулина служили поджелудочная железа крупного рогатого скота, свиней, овец, однако, на такой инсулин у всех больных образуются антитела к этому гормону. Инсулин свиней обладает менее выраженной способностью вызывать образование антител.
Инсулин синтезирован из отдельных аминокислот, но этот метод дорог. Инсулин человека можно получить 2-мя путями: 1) удаляя аминокислоту аланин из В-цепи свиного инсулина и заменяя ее на треонин; 2) с помощью технологии рекомбинантных ДНК, которая успешно применено для получения соматостатина. Для этого встроили ген проинсулин в E. Coli, выделили проинсулин, и получили инсулин протеолизом. Другой способ: синтезировать гены А и В цепей инсулина, встроить их в различные клетки E. Coli, получить эти цепи и связать их дисульфидными мостиками. Инсулин, полученный вторым способом, и свиной инсулин с успехом используются для лечения.
Регуляция секреции
Различают базальную и стимулированную секрецию инсулина. Базальная секреция инсулина – это секреция его в кровь в отсутствие стимулов, т.е. натощак, при голодании и при уровне глюкозы крови 3,2-5,5 ммоль/л (54-100 мг/дл). Хотя считается, что такая концентрация глюкозы в плазме крови не стимулирует секрецию инсулина, она необходима для поддержания базальной секреции инсулина. Базальный уровень инсулина не превышает 69 пмоль/л (0,4 нг/мл).
Механизмы, ответственные за секрецию инсулина и поддержание его уровня в базальном состоянии (натощак, при голодании и в промежутках между приемами пищи), не полностью понятны.
Стимулированная секреция инсулина происходит в ответ на экзогенные стимулы. Наиболее мощным стимулятором секреции инсулина является глюкоза. Механизм ее действия точно не установлен. Полагают, что в В-клетках секрецию инсулина стимулируют метаболиты глюкозы, которые действуют через кальций, способствуя выходу последнего из микротрубочек клеток. Кальций стимулирует выход С-пептида и инсулина из аппарата Гольджи и секрецию их в плазму крови путем экзоцитоза. Возможно, что действие глюкозы в В-клетках на кальций связано с активацией ею циклическим аденозинмонофосфатом (цАМФ).
Помимо глюкозы, секрецию инсулина, но в меньшей степени, стимулируют аминокислоты, в частности лейцин, вагусные влияния, препараты сульфонилмочевины.
Стимулированная секреция инсулина подразделяется на две фазы: начальную (короткую) и медленную (длительную). Считается, что ранняя фаза стимулированной секреции инсулин, является следствием выхода в кровь запасенного в гранулах инсулина, а вторая (медленная) фаза — это секреция вновь синтезированного инсулина.
Угнетают продукцию инсулина симпатические воздействия, катехоламинаы, гормон роста.
У здоровых лиц уровень стимулированного (постпищевого) инсулина в плазме крови редко превышает 690 пмоль/л (100 нг/мл).
В В-клетках содержится островковый амилоид РР — амилин. Это полипептид из 37 аминокислот, который вместе с инсулином запасается в В-клетках островков. На одну молекулу амилоида РР приходится 100 молекул инсулина. Амилин (амилоид РР) секретируется с инсулином в ответ на стимуляцию глюкозой и другие стимуляторы В-клеток. Функция амилина не установлена. Он в избытке синтезируется и откладывается в островках у больных диабетом тип 2 при длительном его течении. Амилин — нерастворимый фибриллярный белок. Оказалось, что у старых людей без диабета, этот белок также накапливается в островках, но в меньшем количестве, чем у лиц с диабетом. Считают, что большое количество амилина у больных диабетом тип 2 является следствием длительной и значительной стимуляции В-клеток глюкозой, а не следствием прямого дефекта гена амилина.
Механизм действия инсулина
Действие инсулина начинается со связывания его со специфическим для него рецептором на плазматической стороне мембраны клеток. Рецептор к инсулину является гликопротеином и состоит из двух субъединиц: большей – α-субъединица и меньшей β-субъединицы. α-Субъединица располагается на наружной поверхности клеточной мембраны и связывает молекулу инсулина, а β-субъединицы преимущественно находится в цитоплазме и содержит тирозинкиназу, которая активируется в ходе связывания инсулина α-субъединицей. В результате происходит аутофосфоризация β-субъединицы и далее каскад энзимных реакций, приводящих к развитию эффектов инсулина.
Чувствительность инсулиновых рецепторов к инсулину, главным образен β-субъединицы зависит от уровня инсулина в плазме крови. При хроническом избытке инсулина (сахарный диабет тип 2, ожирение, хроническая передозировка инсулина, большое количество углеводов в пище) чувствительность к инсулину снижается, развивается так называемый эффект даун-регуляции. Напротив, при низком уровне инсулина в плазме крови (натощак, при голодании, физических упражнениях) чувствительность рецепторов инсулина увеличивается и повышается связывание ими инсулина. Уменьшает связывание инсулина его рецепторами кортизол. Действие кортизола осуществляется как прямо на рецептор к инсулину, так и путем увеличения секреции инсулина в кровоток.
Физиологическая роль инсулина
Главная функция инсулина состоит в утилизации и запасании в клетки тела энергетических и пластических веществ (гликоген, триглицериды, белок, холестерин) из поступающей в организм пищи. Инсулин увеличивает примерно в 20 раз проницаемость клеток мишений для глюкозы и ряда аминокислот, способствует их утилизации. Суммарный эффект – снижение глюкозы в крови.
Инсулин обладает паракринным и эндокринным эффектами:
1. Эндокринный эффект – это действие инсулина на отдаленные от места его образования органы и ткани. Прямо или косвенно инсулин оказывает действие на все ткани. Основными инсулинозависимыми тканями являются печень, жировая и мышечная ткани. Центральная и периферическая нервные системы инсулинонезависимые. Они утилизируют глюкозу без участия инсулина, но являются полностью глюкозозависимыми.
Таблица 6.2.