

5.1. Мозговой слой надпочечников
Мозговой слой находится в центральной части надпочечника и составляет 10% его массы. Мозговое вещество образует сероватую «сердцевину» железы и состоит из групп клеток, окруженных кровеносными сосудами.
Мозговой и корковый слои являются полностью разными структурами. Корковый слой имеет эктодермальное происхождение, а мозговой происходит от первичного нервного гребешка и в конечном итоге является высоко специализированной частью симпатической нервной системы.
М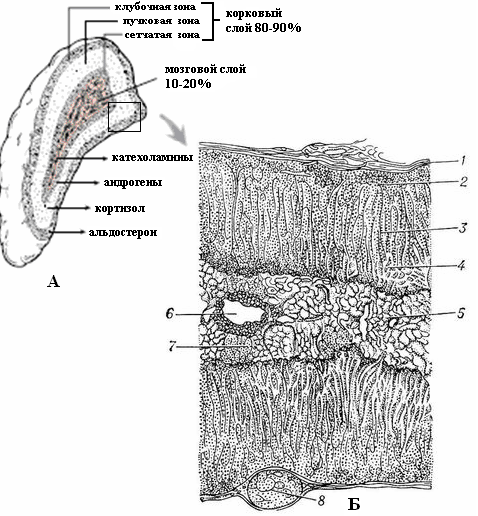 озговое
вещество отделено от коркового вещества
тонкой прерывистой прослойкой
соединительной ткани (рис. 5.1.1). У мозгового
и коркового вещества надпочечника
имеется общее кровоснабжение. Артерии,
входящие в надпочечник, разветвляются
на артериолы, образующие густую
субкапсулярную сеть, от которой отходят
капилляры, снабжающие кровью кору. Их
эндотелий фенестрирован, что облегчает
поступление кортикальных стероидных
гормонов из клеток коркового слоя в ток
крови. Из сетчатой зоны капилляры
вступают в мозговую часть, где принимают
вид синусоидов и сливаются в венулы,
которые переходят в венозное сплетение
мозгового вещества.
озговое
вещество отделено от коркового вещества
тонкой прерывистой прослойкой
соединительной ткани (рис. 5.1.1). У мозгового
и коркового вещества надпочечника
имеется общее кровоснабжение. Артерии,
входящие в надпочечник, разветвляются
на артериолы, образующие густую
субкапсулярную сеть, от которой отходят
капилляры, снабжающие кровью кору. Их
эндотелий фенестрирован, что облегчает
поступление кортикальных стероидных
гормонов из клеток коркового слоя в ток
крови. Из сетчатой зоны капилляры
вступают в мозговую часть, где принимают
вид синусоидов и сливаются в венулы,
которые переходят в венозное сплетение
мозгового вещества.
Рис. 5.1.1. Макро- (А) и микроскопическое (Б) строение надпочечника (секреция соответствующих гормонов):
1 – капсула;
2 – клубочковая зона коры;
3 – пучковая зона коры;
4 – сетчатая зона коры;
5 – мозговое вещество;
6 – вена мозгового вещества;
7 и 8 – смещенные участки коркового вещества.
Железистые клетки мозгового вещества получили название хромаффинных или феохромных. Хромаффинные клетки содержат гранулы с электроплотным содержимым, которое с бихроматом калия дает хромаффинную реакцию.
Хромаффинные клетки содержат многочисленные митохондрии, комплекс Гольджи, элементы гранулярной эндоплазматической сети, многочисленные электроноплотные гранулы, содержащие преимущественно норадреналин и/или адреналин (по этому признаку хромаффинные клетки подразделяются на две субпопуляции), а также АТФ, энкефалины и хромогранины (рис. 5.1.2). Кроме катехоламинов, гранулы содержат липиды, нуклеотиды (АТФ), белки, ионы Са2+ и Mg2+. Катехоламины хромаффинных клеток содержатся в гранулах, одетых липоидной оболочкой и заполненных мелкой зернистостью. Эти гранулы являются специфическими органоидами хромаффинной ткани, на которых протекает биосинтез катехоламиновых секреторных продуктов.
В гранулах мозгового слоя надпочечников содержится 80% адреналина и 20% норадреналина. Норадреналиновые клетки расположены в центре, а адреналиновые – по периферии мозгового слоя надпочечников.
Относительное содержание этих двух гормонов варьирует в зависимости от вида и от стадии развития организма. У большинства млекопитающих норадреналин преобладает до рождения и на первых стадиях неонатального периода, однако, с возрастом в количественном отношении начинает преобладать адреналин. Скорость наступления этого сдвига у различных видов различна: у человека он происходит не ранее трехлетнего возраста.
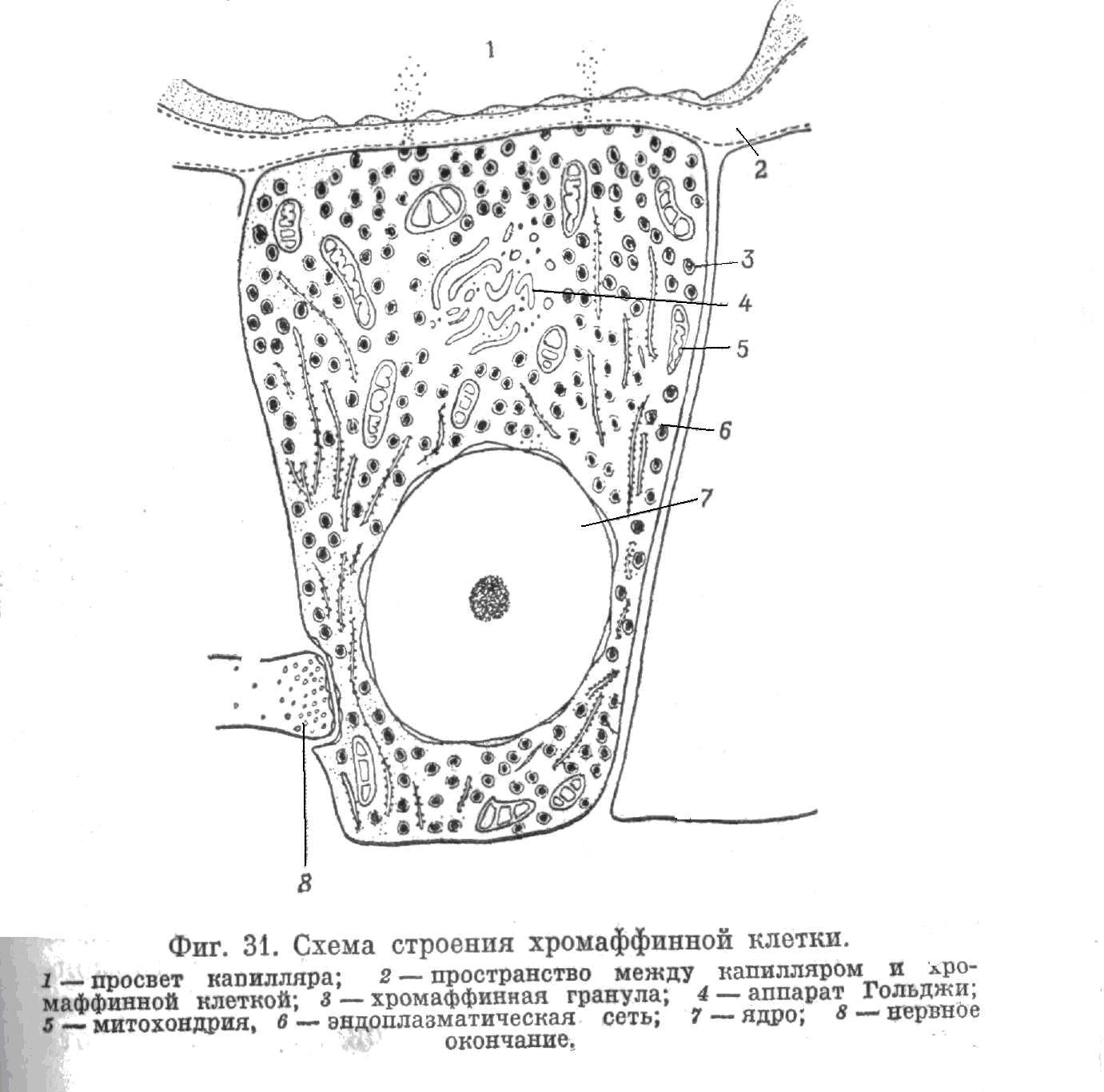
Рис.5.1.2. Строение хромаффинной клетки:
1 – просвет капилляра;
2 – пространство между капилляром и хромаффинной клеткой;
3 – хромаффинная гранула;
4 – аппарат Гольджи;
5 – митохондрия;
6 – эндоплазматическая сеть;
7 – ядро;
8 – нервное окончание.
Хромаффинные клетки — основной клеточный элемент не только мозговой части надпочечников. Мелкие скопления и одиночные хромаффинные клетки находят также в сердце, почках, симпатических ганглиях. Скопление вненадпочечниковой хромаффинной ткани на передней поверхности аорты настолько велико и постоянно, что оно получило даже особое название – орган Цукеркандля. Это так называемая экстрамедуллярная (вненадпочечниковая) хромаффинная ткань (рис. 5.1.3).
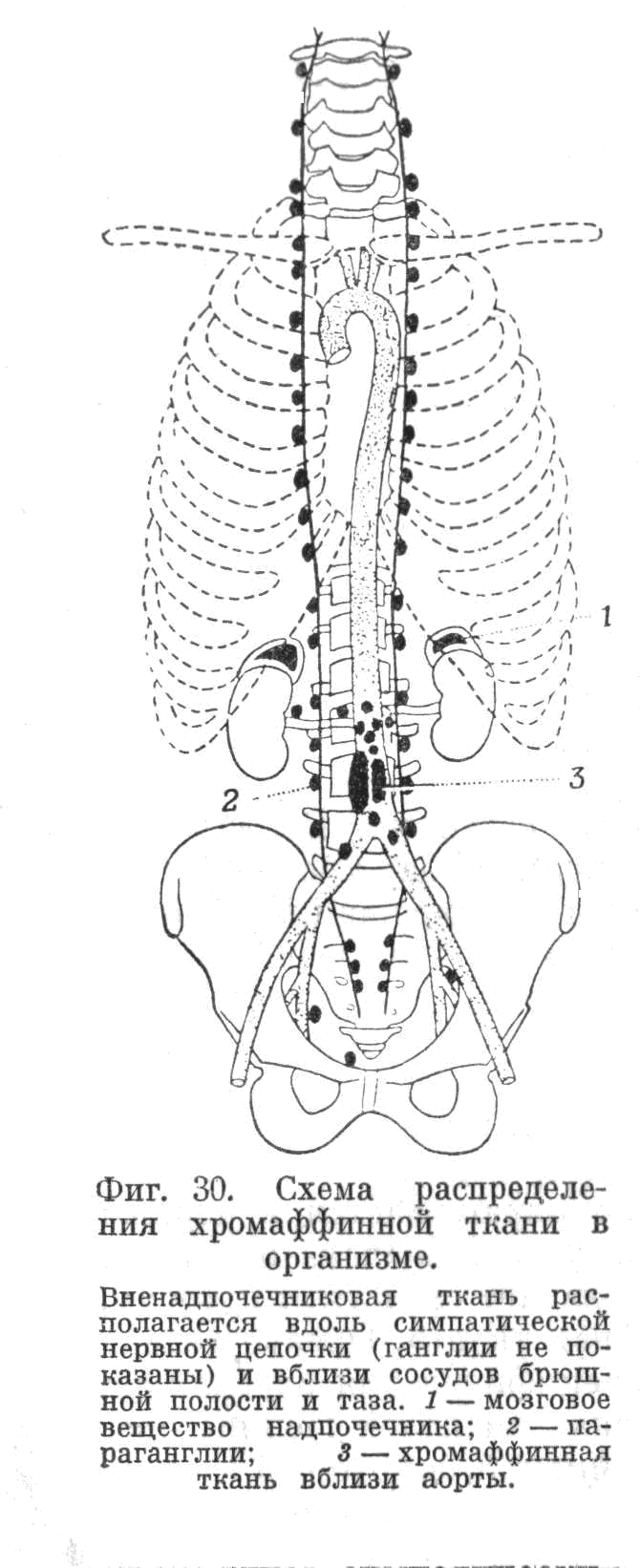
Рис. 5.1.3. Схема распределения хромаффинной
ткани в организме.
Вненадпочечниковая («экстрамедуллярная») ткань
располагается вдоль симпатической нервной
цепочки (ганглии не показаны) и вблизи сосудов
брюшной полости и таза.
1 – мозговое вещество надпочечников;
2 – параганглии;
3 – хромаффинная ткань вблизи аорты.
Все хромаффинные клетки имеют общее происхождение в эмбриогенезе, возникая из нейробластов (первичных нервных клеток), расположенных главным образом в грудном отделе центральной нервной системы. Эти первичные нервные клетки мигрируют из места своего возникновения по ходу симпатических нервов. Экстрамедуллярная хромаффинная ткань секретирует, главным образом, норадренали, напоминая в этом отношении постганглионарные нейроны симпатической нервной системы.
Функция хромаффинной клетки регулируется симпатической нервной системой. Морфологическая связь нервных волокон с хромаффинной клеткой различна. Нервные окончания могут просто подходить к поверхности клетки, а могут и вдавливаться в клеточную мембрану так, что оказывается более или менее окруженными ею. Однако независимо от их взаимного расположения между мембраной нервного окончания и мембраной клетки всегда есть щель шириной ≈ 150Å. Наружная пограничная мембрана этих двух компонентов на месте сближения утолщена. Таким образом, здесь формируется синапс. В пресинаптической части, образованной аксоном, расположены синаптические пузырьки, в которой находится ацетилхолин. В момент передачи возбуждения от симпатического волокна хромаффинным клеткам (например, при раздражении чревного нерва) в синаптическую щель 1) выделяется ацетилхолин, который диффундирует через узкую синаптическую щель к мембране хромаффинной клетки;
2) далее ацетилхолин взаимодействует со специфическими рецепторными участками мембраны хромаффинной клетки и активирует ее;
3) через активированную мембрану в клетку мигрируют ионы кальция, который взаимодействует с определенными реактивными участками цитоплазмы;
4) этот процесс стимулирует выделение из клетки катехоламинов и АТФ.
В связи с этим хромаффинную ткань можно рассматривать как модифицированное периферическое звено симпатической нервной системы, осуществляющее свои эффекты гуморально. Мозговой слой надпочечников и симпатическая нервная система функционируют в тесной связи друг с другом и представляют единую систему регуляции, обычно обозначаемую симпатоадреналовой.
В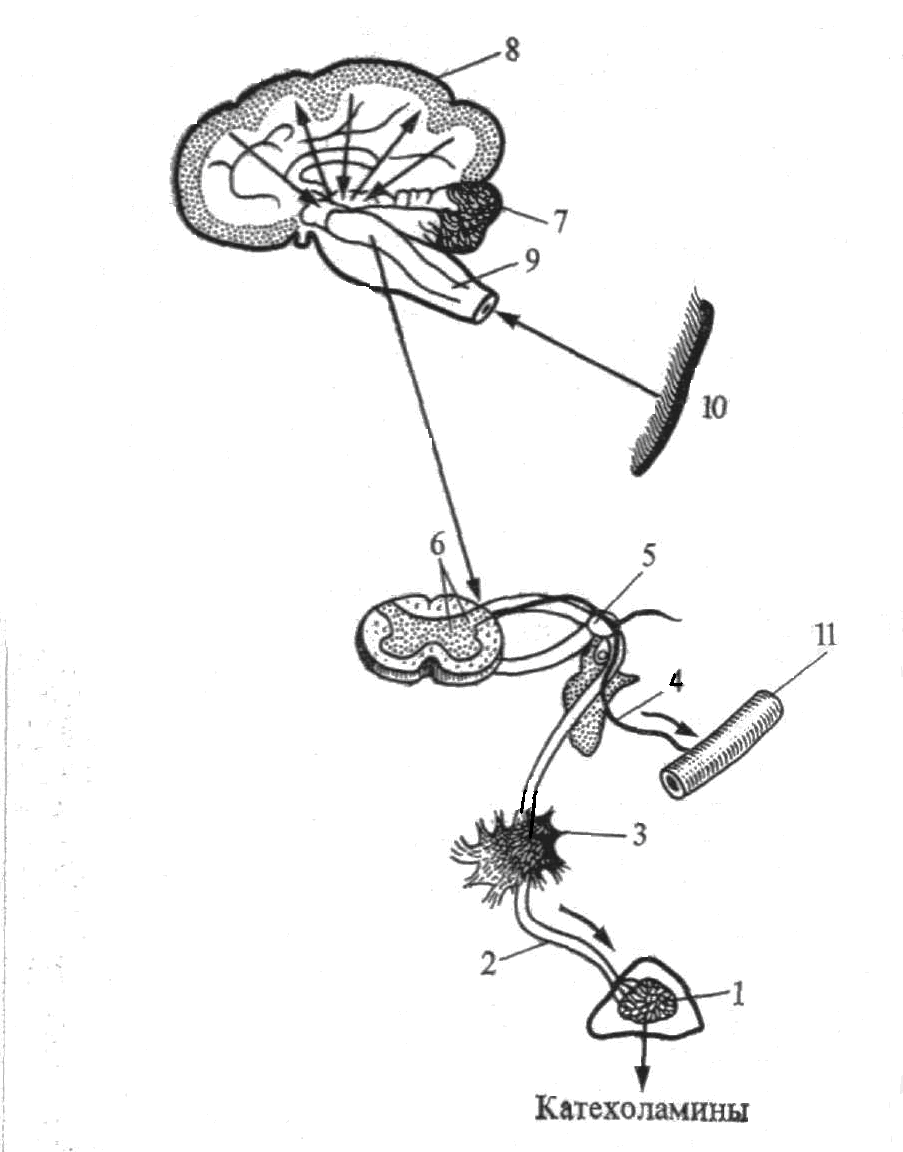 период интенсивной
симпатической стимуляции (холод,
чрезмерная физическая активность)
мозговой слой надпочечников прогрессивно
увеличивает секрецию катехоламинов
(рис. 5.1.4.). В других ситуациях симпатическая
нервная система и мозговой слой
надпочечников стимулируются независимо
друг от друга. Например, вертикальное
положение тела стимулирует симпатическую
нервную систему, а гипогликемия
стимулирует только мозговой слой
надпочечников. В ситуациях, когда
симпатическая
нервная система подавлена, мозговой
слой надпочечников снабжает организм
катехоламинами и поддерживает его
жизненно важные функции.
период интенсивной
симпатической стимуляции (холод,
чрезмерная физическая активность)
мозговой слой надпочечников прогрессивно
увеличивает секрецию катехоламинов
(рис. 5.1.4.). В других ситуациях симпатическая
нервная система и мозговой слой
надпочечников стимулируются независимо
друг от друга. Например, вертикальное
положение тела стимулирует симпатическую
нервную систему, а гипогликемия
стимулирует только мозговой слой
надпочечников. В ситуациях, когда
симпатическая
нервная система подавлена, мозговой
слой надпочечников снабжает организм
катехоламинами и поддерживает его
жизненно важные функции.
Рис. 5.1.4. Схема нервной регуляции функций мозгового слоя надпочечников:
1 – мозговой слой надпочечника;
2 – ветвь большого чревного нерва;
3 – солнечное сплетение;
4 − вазомоторный нерв;
5 − симпатический ганглий;
6 – спинномозговой симпатический центр;
7 – симпатические центры гипоталамуса;
8 – кора больших полушарий;
9 – ретикулярная формация;
10 – рецептивное поле;
11 − сосуд.
Биосинтез катехоламинов
Как надпочечниковая («медуллярная»), так и вненадпочечниковая («экстрамедуллярная») хромаффинная ткань выделяют гормоны адреналин и норадреналин, объединенных под общим названием катехоламины. Адреналин синтезируется только в надпочечниках; норадреналин и дофамин образуются также в параганглиях и многочисленных нейронах симпатической нервной системы. Эти эндокринные клетки иннервируются волокнами симпатической нервной системы.
Таким образом, синтезированные в нейронах катехоламины являются нейротрансмиттерами, которые опосредуют функцию ЦНС и симпатической нервной системы.
Катехоламины синтезируются из аминокислоты тирозин путем ряда превращений, регулируемых определенными энзимами, который в мозговом слое надпочечников в хромаффинных клетках под влиянием энзима тирозингидроксилазы превращается в дегидрооксифенилаланин (ДОФА) (рис. 5.1.5). Далее ДОФА под влиянием энзима декарбоксилазы превращается в дофамин, а последний гидроксилируется энзимом дофаминбетагидроксилазой в норадреналин. В окончаниях периферических симпатических нервов синтез дофамина и норадреналина – идет тем же путем и под влиянием тех же энзимов, что и в мозговом слое надпочечников и в мозге. В мозговом слое надпочечников норадреналин под влиянием энзима N-метилтрансферазы превращается в адреналин. В симпатической нервной системе (в основном, в постганглионарных окончаниях симпатических нервов) адреналин не образуется, так как энзим, необходимый для его образования (N-метилтрансфераза) имеется только в мозговом слое надпочечников.
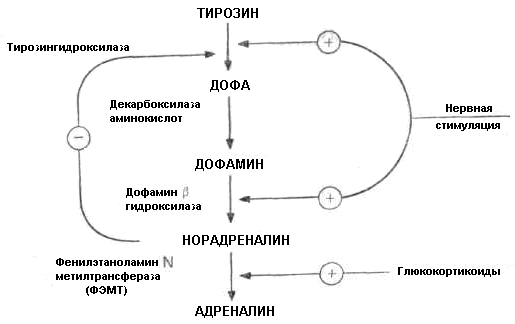
Рис. 5.1.5. Регуляция синтеза катехоламинов в мозговом слое надпочечников.
При увеличении образования дофамина декарбоксилаза тормозит активность тирозингидроксилазы и синтез катехоламинов уменьшается. При уменьшении образования дофамина активность тирозингидроксилазы возрастает и синтез катехоламинов увеличивается. Кроме того, синтез адреналина регулируется кортизолом, который поступает в мозговой слой из кортикомедуллярного венозного синуса. Кортизол активирует энзим N-метил-трансферазу, превращающую норадреналин в адреналин.
Секреция катехоламинов осуществляется путем экзоцитоза; при этом содержание гранул “изливается” во внеклеточное пространство.
Высвобождение катехоламинов как из мозгового слоя надпочечников, так и из окончаний симпатической нервной системы происходит под влиянием таких физиологических стимуляторов, как стресс, физическая и психическая нагрузка, повышение уровня инсулина в крови, гипогликемия, гипотония и др. Высвобождение катехоламинов происходит при участии ионов Са2+, который поступает в клетку или в окончания симпатической нервной системы. Поступающие в кровь катехоламины достигают периферических тканей, где накапливаются или метаболизируются прямо пропорционально симпатической иннервации тканей.
Механизм действия катехоламинов
Адреналин и норадреналин вызывают эффекты, которые различны во многих отношениях.
Эффекты катехоламинов при их воздействии, например, на сосуды различных органов, различаются (сосуды мышц – расширяются, кишечника – суживаются, коронарный кровоток под влиянии адреналина снижается, норадреналина - возрастает). Для объяснения этих эффектов была предложена теория клеточных рецепторов (Алквист, 1948). Согласно Алквисту существует 2 типа рецепторов, реагирующих с катехоламинами – α и β.
α-адренэргическое действие охватывает такие быстрые эффекты, как вазоконстрикцию, сокращение капсул селезенки, матки, семявыносящих протоков, а также торможение гладкой мускулатуры ЖКТ, мочевого пузыря. Через α-адренорецепторы действует, в основном, норадреналин, адреналин значительно слабее (табл. 5.1.1).
Таблица 5.1.1.