

4.2. Гормоны нейрогипофиза
Задняя доля гипофиза или нейрогипофиз образуется в эмбриогенезе как выпячивание вентрального гипоталамуса. В нейрогипофизе локализованы веретенообразные клетки – питуициды и аксоны гипоталамических нейронов.
Нейрогипофиз не образует, а лишь накапливает и выделяет нейрогормоны супраоптического и паравентрикулярного ядер гипоталамуса, т.е. он не является секреторным и органом. Эти данные были получены с помощью метода радиоавтографии.
Еще в 1928 г. с помощью классических методов окрашивания в нейронах – супраоптического и паравентрикулярного ядер – были обнаружены секреторные гранулы. Аксоны этих нейронов, через гипофизарную ножку идут к задней доле гипофиза (рис. 4.1). В этих аксонах были обнаружены секреторные капельки; такие же капельки, как было явственно видно, накапливаются и в задней доле гипофиза. На основании этих фактов, выявленных путем гистологических исследований, возникла мысль о том, что нейросекрет из нейронов гипоталамуса проходит по аксонам в нейрогипофиз, где из нервных окончаний высвобождаются полипептидные гормоны окситоцин и вазопрессин. Методом радиоавтографии (с использованием аминокислот, меченных S35) была изучена динамика этого процесса. Вазопрессин и окситоцин – полипептиды, и в числе составляющих их аминокислот имеется серосодержащая аминокислота цистеин. Цистеин, меченный S35, вводили в желудочки мозга, и через разные промежутки времени после этого животных забивали. Затем готовили срезы мозга и гипофиза и исследовали их методом радиоавтографии. Изучение радиоавтографов показало, что вскоре после введенпя 835-цистеина радиоактивность появляется в гипоталамусе (рис. 4.2.1) и затем постепенно накапливается в задней доле гипофиза. Запаздывание поступления S35 в заднюю долю гипофиза характеризует время, необходимое для того, чтобы нейросекрет, выработавшийся в нейронах гипоталамуса, переместился по аксонам в гипофиз.
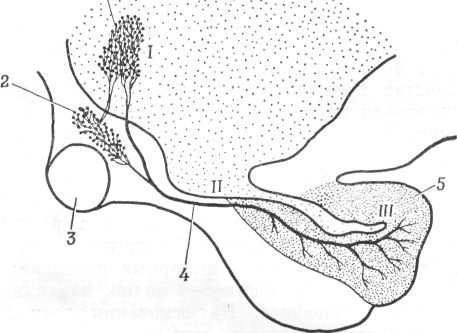
Рис. 4.2.1. Схема радиоавтографического исследования секреции окситоцина и вазопрессипа.
Сагиттальный разрез через гипоталамус и гипофиз; показан нервный тракт, связывающий супраоптическое и паравентрикулярное ядра гипоталамуса и заднюю долю гипофиза, а также распределение и последовательность появления радиоактивности (I-III) в этих отделах мозга после введения в желудочек мозга меченых предшественников окситоцина и вазопрессина.
1 – паравентрикулярное ядро в дне третьего желудочка; 2 – супраоптическое ядро; 3 – перекрест зрительных нервов; 4 – гипоталамо-гипофизарный тракт; 5 – задняя доля гипофиза.
Cупраоптическое ядро гипоталамуса состоит из нейронов, секретирующих вазопрессин (антидиуретический гормон АДГ). Аксоны этих нейронов направляются в заднюю долю гипофиза.
Паравентрикулярное ядро гипоталамуса содержит 8 клеточных зон; образованных нейронами, которые синтезируют различные вещества: АДГ, окситоцин, кортикотропинрелизинт фактор, соматостатин, энкефалин и т.д. Терминали этих нейронов обнаружены не только в нейрогипофизе, но и в срединном возвышении, различных отделах ЦНС.
Миграция секрета (нейрогормонов) по аксонам происходит со скоростью 3 мм в сутки. Они накапливаются в терминалях, а при стимуляции – выделяются в кровь.
Расшифровка структуры окситоцина и вазопрессина и их последующий химический синтез можно считать наиболее важными событиями в современной биохимии. Окситоцин был впервые выделен и синтезирован в 1953 г. Винсентом дю Виньо (США), за что ему была присуждена Нобелевская премия по химии. Оба гормона близки к октапептидам, в которых аминокислоты замыкаются в кольцо за счет S-S связей. Аргинин-вазопрессин обнаружен у человека, лошади, овцы и т.д.; для свиньи и гиппопотама характерен лизин-вазопрессин, содержащий вместо аргинина лизин. Синтезировано множество аналогов пептидов задней доли гипофиза.