

4. Гипофиз
Гипофиз располагается в основании черепа, в выемке решетчатой кости, которая называется «турецкое седло». Гипофиз окружен твердой мозговой оболочкой и сверху его отделяет от других частей мозга диафрагма турецкого седла (рис. 4.1). Она препятствует попаданию в турецкое седло спинномозговой жидкости. К гипофизу через отверстие в диафрагме проходит ножка гипофиза, окруженная кровеносными сосудами. Вес гипофиза колеблется от 600 до 1200мг. Во время беременности вес гипофиза увеличивается за счет гипертрофии лактотрофов и увеличения кровотока.
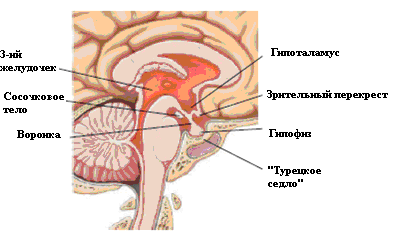
Рис. 4.1. Взаимосвязь гипофиза с гипоталамусом.
Гипофиз состоит из двух долей – большей передней доли и меньшей задней доли. Передняя доля, или аденогипофиз, эктодермального происхождения. Она образуется из эктодермы орофарингса, которая внедряется в турецкое седло. Задняя доля, или нейрогипофиз, образуется путем инвагинации в турецкое седло вентрального отдела гипоталамуса. Задняя доля состоит из аксонов нейронов, расположенных в супраоптических и паравентрикулярных ядрах гипоталамуса. Аксоны этих клеток проходят через ножку гипофиза. В задней доле оканчиваются на капиллярах, которые собираются в вену задней доли, впадающую в кавернозный синус. В задней доле освобождаются в кровоток гормоны, синтезированные в гипоталамусе, – вазопрессин и окситоцин.
Между задней и передней долями гипофиза располагается очень небольшая часть гипофиза, остаток кармана Ратке, которая называется средней долей. У человека она практически гормонально неактивна. Предполагается, что в ней может синтезироваться предшественник АКТГ – меланоцитостимулирующий гормон.
4.1. Строение и функциональная роль аденогипофиза
4.1.1. Клеточное строение аденогипофиза
Структура передней доли гипофиза предоставлена 8 типами клеток, из которых основная секреторная функция присуща хромафильным клеткам 5 групп. Выделяют следующие типы клеток: 1) ацидофильные красные клетки с мелкими гранулами или соматотрофы — вырабатывают соматотропин (СТГ, гормон роста); 2) ацидофильные желтые клетки — с крупными гранулами или лактотрофы — вырабатывают пролактин; 3) базофильные тиреотрофы — вырабатывают тиреотропин (тиреотропный гормон — ТТГ); 4) базофильные гонадотрофы — вырабатывают гонадотропины: фоллитропин (фолликулостимулирующий гормон — ФСГ) и лютропин (лютеинизирующий гормон — ЛГ); 5) базофильные кортикотрофы — вырабатывают кортикотропин (адренокортикотропный гормон — АКТГ). Кроме того, также как и в клетках промежуточной доли, в базофильных кортикотрофах образуются β-эндорфин и меланотропин, поскольку все эти вещества происходят из общей молекулы предшественника липотропинов.
Таким образом, в аденогипофизе синтезируются и секретируются пять основных типов гормонов: кортикотропин, гонадотропины (фоллитропин и лютропин), тиреотропин, пролактин и соматотропин. Первые три из них обеспечивают гипофизарную регуляцию периферических эндокринных желез (коры надпочечников, половых желез и щитовидной железы), т.е. участвуют в реализации гипофизарного пути управления. Для двух других гормонов (соматотропина и пролактина) гипофиз выступает в роли периферической эндокринной железы, поскольку эти гормоны действуют на ткани-мишени (рис. 4.1.1, табл. 4.1).
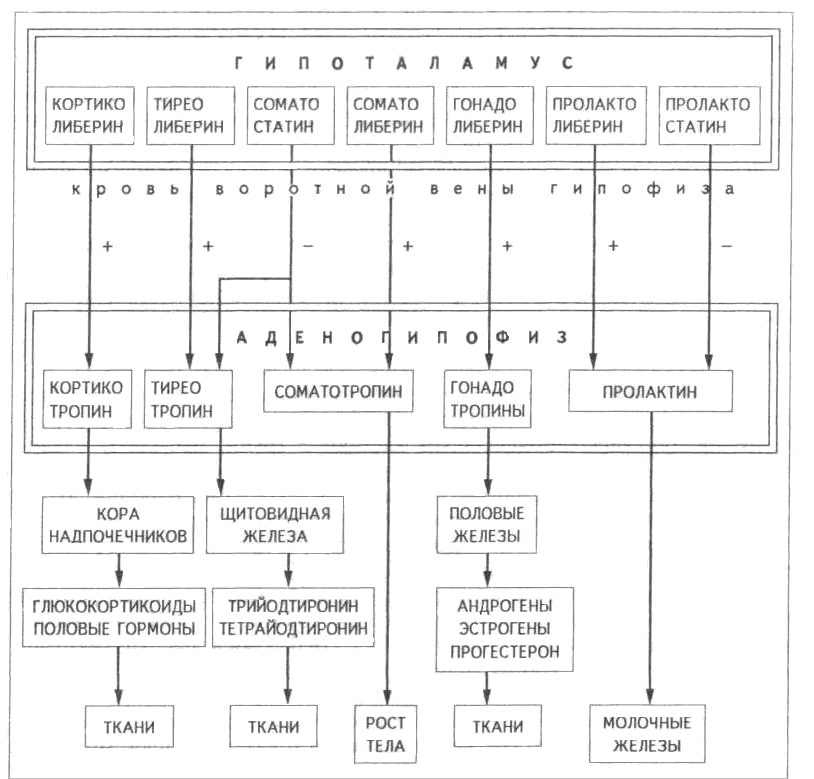
Рис. 4.1.1. Основные гормоны аденогипофиза, регуляция их секреции и мишени.
(+) – стимуляция, (-) – подавление секреции.
Таблица 4.1.
Гормон |
Химическая природа |
Ткань-мишень |
Главные эффекты у млекопитающих |
Регуляция секреции |
Аденокортикотропный гормон (АКТГ) |
Пептид n = 39 |
Кора надпочечников |
Стимулирует синтез и секрецию стероидов корой надпочечников |
Стимулируется кортиколиберином |
Тиреотропный гормон (ТТГ) |
Гликопротеин |
Щитовидная железа |
Усиливает синтез и секрецию тиреоидных гормонов |
Стимулируется тиреолиберином и подавляется тиреоидными гормонами |
Гормон роста (СТГ) |
Полипептид |
Все ткани |
Ростовое действие, синтез белка, стимуляция липолиза, снижение чувствительности ткани к инсулину |
|
Фолликулостимулирующий гормон (ФСГ) |
Гликопротеин |
Семенные канальцы у самцов, фолликулы яичников у самок |
У самцов повышает образование спермы, у самок стимулирует созревание фолликулов |
Стимулируется люлиберином и подавляется ингибином |
Лютеинизирующий гормон (ЛГ) |
Гликопротеин |
Интерстициальные клетки семенников (у самцов) и яичников (у самок) |
Вызывает окончательное созревание фолликулов, секрецию эстрогенов, овуляцию, образование жёлтых тел, секрецию прогестерона у самок; усиливает синтез и секрецию андрогенов у самцов |
Стимулируется люлиберином |
Пролактин |
Полипептид n = 198 |
Молочная железа (альвеолярные клетки) |
Стимулирует синтез белков молока и развитие молочных желез; пробуждает родительский инстинкт у самок |
Находится под постоянным тормозящим действием гипоталамического фактора (пролактостатина); она становится возможной при повышенной концентрации эстрогенов и пониженной секреции пролактастатина |
α-Меланоцитстимулирующий гормон (α-МСГ) |
Олигопептид n = 13 |
Пигментные клетки |
Повышает синтез меланина в меланоцитах и увеличивает размеры меланофоров (вызывает потемнение кожи) |
Подавляется гипоталамическим фактором (меланостатином) |
4.1.2. Регуляция секреции и физиологические эффекты кортикотропина.
Кортикотропин является продуктом расщепления крупного (239 аминокислот) гликопротеина проопиомеланокортина, образующегося базофильными кортикотрофами. Этот белок делится на две части, одна из которых при расщеплении служит источником кортикотропина и меланотропина, а вторая, называемая липотропином, расщепляясь, дает кроме меланотропина морфиноподобный пептид эндорфин, играющий важнейшую роль в антиноцицептивной (антиболевой) системе мозга и в модуляции секреции гормонов аденогипофиза.
Секреция кортикотропина происходит постоянно пульсирующими вспышками с четкой суточной ритмичностью. Наивысшая концентрация гормона в крови отмечается в утренние часы, а наиболее низкая — с 22 до 2 часов ночи. Регуляция секреции представлена прямыми и обратными связями. Прямые связи реализуются кортиколиберином гипоталамуса, а обратные запускаются содержанием в крови гипофиза кортикотропина и уровнем гормона коры надпочечников кортизола в системной циркуляции. Обратная связь имеет отрицательную направленность и замыкается как на уровне гипоталамуса (подавление секреции кортиколиберина), так и гипофиза (торможение секреции кортикотропина). Продукция кортикотропина резко возрастает при действии на организм сильных раздражителей, например, холода, боли, физической нагрузки, эмоций, а также под влиянием гипогликемии (снижение сахара в крови).
Физиологические эффекты кортикотропина делят на надпочечниковые и вненадпочечниковые. Надпочечниковое действие гормона является основным и заключается в стимуляции (через системы аденилатциклаза-цАМФ и Са++) клеток пучковой зоны коры надпочечников, секретирующей глюкокортикоиды (кортизол и кортикостерон) (рис. 4.1.2, 4.1.3). Значительно меньший эффект кортикотропин оказывает на клетки клубочковой и пучковой зон коры надпочечников, т.е. на продукцию минералокортикоидов и половых стероидов.
Синтез минералокортикоидов регулируется ренинангиотензионовой системой и уровнями натрия и калия в сыворотке крови.
Под влиянием кортикотропина усиливается стероидогенез (синтез гормонов), за счет повышения образования и активации транскрипции генов, что при избытке гормона вызывает гипертрофию и гиперплазию коры надпочечников.
Вненадпочечниковое действие кортикотропина заключается в следующих эффектах: 1) липолитическое действие на жировую ткань, 2) повышение секреции инсулина и соматотропина, 3) гипогликемия из-за стимуляции секреции инсулина, 4) повышенное отложение меланина с гиперпигментацией из-за родства молекулы гормона с меланотропином.
Избыток кортикотропина сопровождается развитием гиперкортицизма с преимущественным увеличением секреции надпочечниками кортизола и носит название "болезнь Иценко- Кушинга". Основные проявления типичны для избытка глюкокортикоидов. Дефицит кортикотропина ведет к недостаточности глюкокортикоидов с выраженными метаболическими сдвигами и снижением устойчивости организма к влияниям среды.
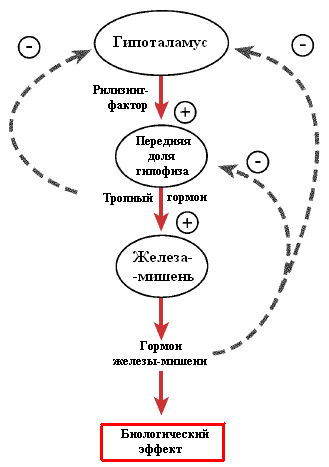
Рис. 4.1.2. Прямая и обратная связи регуляции секреции тропных гормонов аденогипофиза.
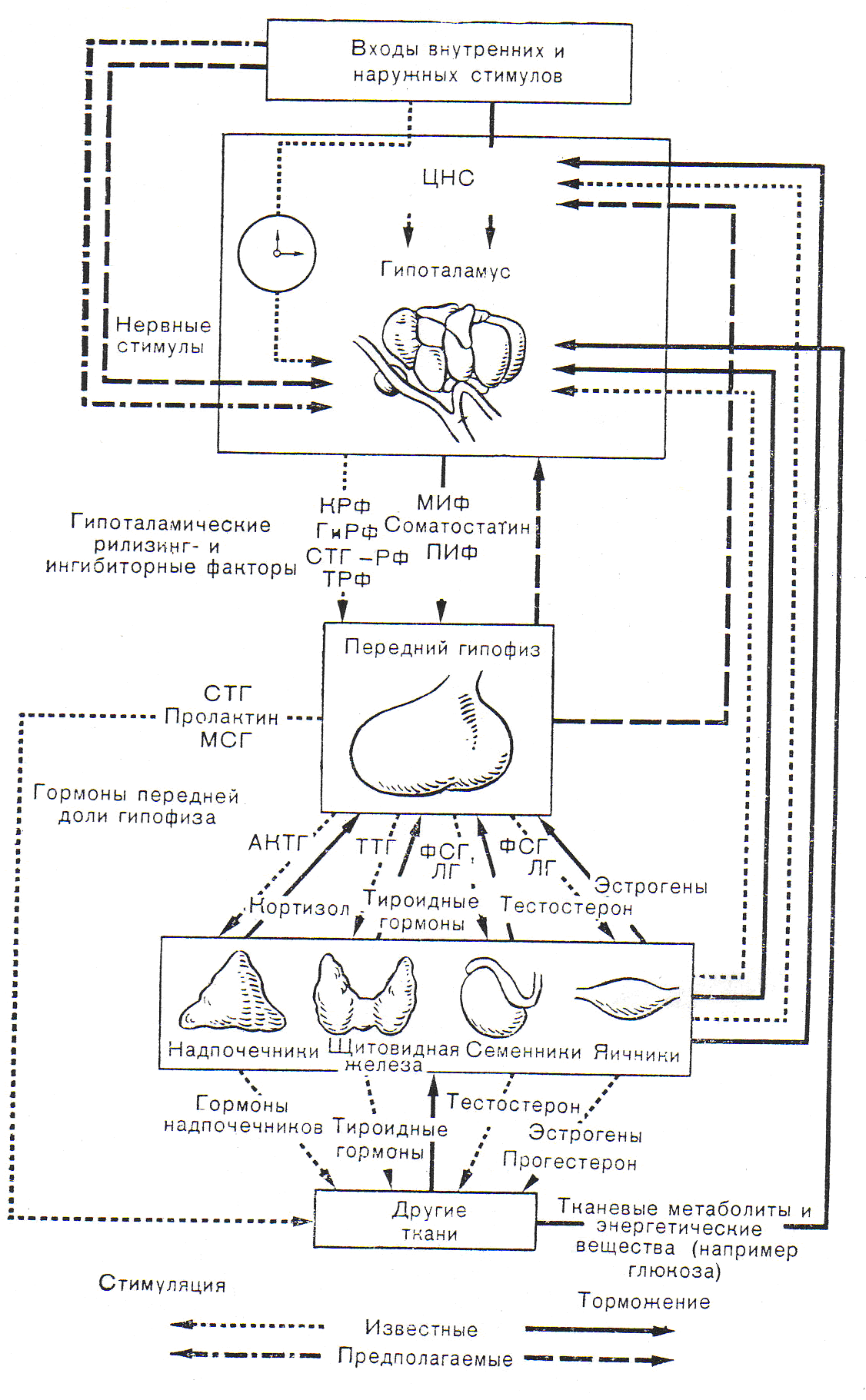
Рис. 4.1.3. Схематическое изображение основных петель обратной связи в нейроэндокринной системе, отражающих взаимодействие ее компонентов.