

Мышечное сокращение
Сокращение мышцы происходит при поступлении по аксонам двигательных нейронов к нервно-мышечным синапсам волны возбуждения в виде нервных импульсов (ПД нервных волокон). Это непрямое сокращение (опосредованное нервно-мышечной синаптической передачей). Возможно и прямое сокращение мышцы. Под ним понимают сокращение групп МВ (мышечные подёргивания, фибрилляции), происходящее при возбуждении любого звена последовательности событий после секреции нейромедиатора из терминалей аксона в нервно-мышечном синапсе. Последовательность этих событий такова: (1) деполяризация постсинаптической мембраны и генерация ПД (2) распространение ПД по плазмолемме МВ (3) передача сигнала в триадах на саркоплазматический ретикулум (4) выброс Ca2+ из саркоплазматического ретикулума (5) связывание Ca2+ тропонином С тонких нитей (6) взаимодействие тонких и толстых нитей (формирование мостиков), появление тянущего усилия и скольжение нитей относительно друг друга (7) Цикл взаимодействия нитей (8) укорочение саркомеров и сокращение МВ (9) расслабление. Позиции 1–4 рассмотрены выше (см. рис. 7–4 и 7–5 в книге и сопровождающий их текст), а этапы 2–4 представлены на рис. 7–10.

Рис. 7–10. Распространение потенциала действия по сарколемме мышечного волокна и выброс ионов кальция из цистерн саркоплазматического ретикулума
1. Деполяризация постсинаптической мембраны и генерация пд рассмотрены выше и в главе 6.
2. Плазмолемма и потенциал действия. Локальная деполяризация постсинаптической мембраны приводит к генерации потенциала действия, быстро распространяющегося по всей плазмолемме мышечного волокна (включая Т-трубочки).
Электромиография — важный диагностический метод — позволяет регистрировать характеристики потенциалов действия.
Миотония. Уменьшение Cl—-проводимости плазмолеммы ведёт к электрической нестабильности мембраны МВ и к развитию миотонии (например, болезни Томсена).
3. Триады и передача сигнала на саркоплазматическую сеть. Волна деполяризации по Т-трубочкам проникает до триад. В области триад мембрана Т–трубочек в составе потенциалозависимого кальциевого канала содержит рецепторы дигидропиридина. Деполяризация мембраны Т–трубочек вызывает в структуре рецепторов дигидропиридина конформационные изменения, передающиеся на рецепторы рианодина терминальных цистерн саркоплазматического ретикулума.
Злокачественная гипертермия при наркозе (особенно при использовании тиопентала и галотана) — редкое осложнение (смертность до 70%) при хирургическом вмешательстве. Температура тела быстро поднимается до 43 °С и выше, происходит генерализованный распад мышц (рабдомиолиз). В части случаев найдена мутация гена рианодинового рецептора скелетномышечного типа.
4. Саркоплазматический ретикулум и выброс Ca2+. Активация рецепторов рианодина (Ca2+‑канал) приводит к открытию Ca2+‑каналов, Ca2+ из кальциевых депо поступает в саркоплазму; концентрация Ca2+ в саркоплазме достигает значений, достаточных для связывания этого двухвалентного катиона с тропонином С тонких нитей.
5. Связывание Ca2+ тонкими нитями. В покое взаимодействие тонких и толстых нитей невозможно, т.к. миозин-связывающие участки F‑актина заблокированы тропомиозином. При высокой концентрации Ca2+ эти ионы связываются с тропонином C и вызывают конформационные изменения тропомиозина, приводящие к разблокированию миозин-связывающих участков (рис. 7–11).
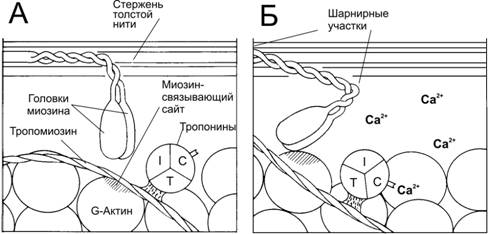
Рис. 7–11. Ca2+–зависимый механизм регуляции взаимодействия актина с миозином [11]. В покое миозин-связывающие участки тонкой нити заняты тропомиозином. При сокращении ионы Ca2+ связываются с тропонином С, а тропомиозин открывает миозин-связывающие участки. Головки миозина присоединяются к тонкой нити и вызывают её смещение относительно толстой нити.
6. Взаимодействие тонких и толстых нитей. В результате разблокирования миозин-связывающих участков молекул актина головки миозина, несущие продукты гидролиза АТФ (АДФ + Фн), присоединяются к тонкой нити и изменяют свою конформацию, создавая тянущее усилие: — тонкие нити начинают скользить между толстыми (рис. 7–12). За счёт шарнирного участка в области шейки миозина происходит гребковое движение, продвигающее тонкую нить к центру сaркомера. В результате происходит скольжение тонких нитей относительно толстых. Затем головка миозина связывается с молекулой АТФ, что приводит к отделению миозина от актина. Последующий гидролиз АТФ восстанавливает конформированную молекулу миозина, готовую вступить в новый цикл. Такая модель скользящих нитей была предложена Хью Хаксли.

Рис. 7–12. Взаимодействие головки миозина с тонкой нитью и появление тянущего усилия