

1.7.16. Мімівірус - недостаюча ланка між вірусами і бактеріями або принципово нова форма життя?
Мімівірус (група dsDNA) — рід вірусів, включає єдиний відомий вид Acanthamoeba polyphaga mimivirus (APMV). APMV зазвичай називають просто мімівірусом. Цей вірус відрізняється найбільшим діаметром капсиду з усіх відомих вірусів, а також, в порівнянні з іншими вірусами, об'ємнішим (більше 1,2 мільйонів пар нуклеотидів) і складноструктурованим геномом. У світлі нестачі точних даних про природу цього вірусу, його відкриття викликало великий інтерес у наукових колах. Було висловлено припущення, що мімівірус є недостаючою ланкою між вірусами і бактеріями. Радикальніша точка зору говорить про те, що мімівірус є принципово новою формою життя, що не відноситься до вірусів або бактерій.
APMV був випадково знайдений в 1992 році, під час дослідження легіонельозу, в амебі Acanthamoeba polyphaga, на честь якої він і був названий.
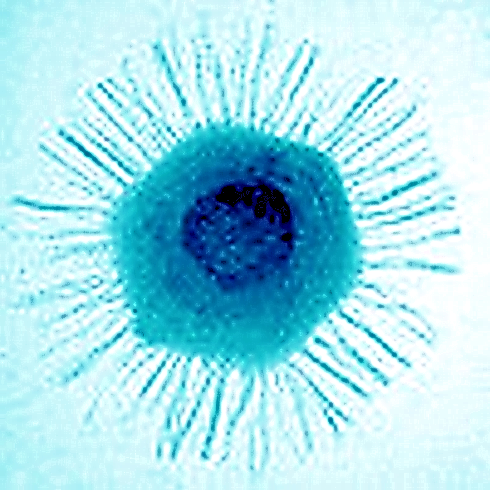
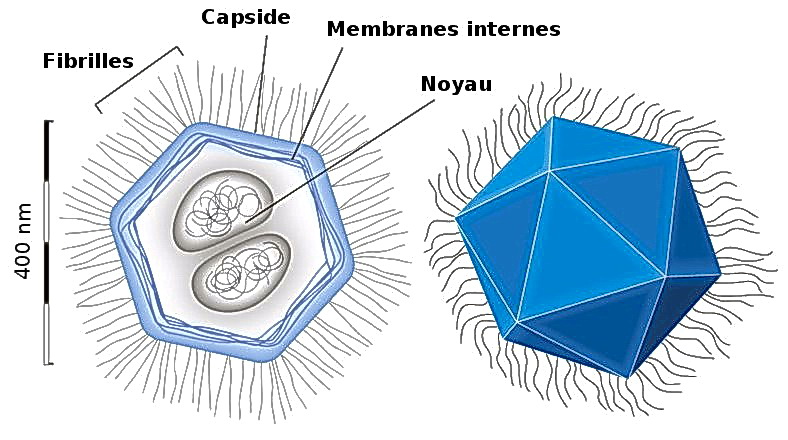
Рис. 26. Мімівірус. Електрона мікрофотографія та схема будови
Вірус був виявлений в препараті, забарвленому по Граму, і внаслідок цього був помилково прийнятий за грампозитивну бактерію. Організм був названий "Bradfordcoccus" на честь району в якому була знайдена амеба (Бредфорд, Англия). У 2003 році дослідники з Universite de la Mediterranee (Марсель, Франція) опублікували статтю в журналі "Science" в якій описали цей мікроорганізм як вірус. Існує гіпотеза, що мімівірус може викликати у людей деякі форми пневмонії. Досі були знайдені лише непрямі свідчення на користь цієї гіпотези у вигляді антитіл до вірусу, виявлених у пацієнтів тих, що страждають пневмонією
Ряд білків мімівіруса, що беруть участь в реплікації генома, були гомологічними білкам інших великих ядерно-цитоплазматичних ДНК вірусів (поксвіруси, ірідовіруси, фікоднавіруси), що може вказувати на їх спільне походження. Проте велика кількість мімівірусних білків не виявляє схожість ні з одним відомим нині білком. Крім того, геном мімівіруса кодує значну кількість білків, подібних до ядерних і бактеріальних. Мабуть ці гени були придбані мімівірусом повторно і походять з геномів хазяїв вірусу і їх паразитів.
Мімівірус, маючи капсид діаметром 400 нм з численними 100-нанометровими білковими нитками на ньому, є найбільшим відомим на сьогодні вірусом. У науковій літературі приведені розміри віріона від 400 нм до 800 нм, залежно від того, чи заміряється діаметр капсиду або загальна довжина вірусу в подовжній осі. У електронному мікроскопі можна спостерігати гексагональну форму капсиду, що вказує на ікосаедральну симетрію білкових структур капсиду. У мімівіруса не спостерігається зовнішньої оболонки, що вказує на те, що мімівірус покидає заражену клітину не шляхом екзоцитозу (відбруньковування).
Мімівірус має багато особливостей будови, характерних і для інших вірусів своєї групи. Центральна частина вірусу – ДНК, виглядає під електронним мікроскопом як електронно-щільна область.
З очищених віріонів були виділені декілька різних мРНК, що кодують ДНК-полимеразу, білки капсиду і фактори транскрипції, близькі до TFII. Також були знайдені мРНК, що кодують аміноацил тРНК синтетазу, і 4 неідентифікованих молекули мРНК, специфічних для мімівіруса. Ці мРНК можуть бути трансльовані без експресії вірусних генів.
Геном мімівіруса складається з лінійної молекули ДНК, що містить близько 1,2 млн пар основ. Це найбільший геном серед усіх відомих науці вірусів, він в два рази довше, ніж наступний за розміром геном міовіруса Bacillus phage G. Крім того, мімівірус володіє більшим об'ємом генетичної інформації ніж мінімум 30 організмів, що мають клітинну будову.
На додаток до унікального для вірусів розміру генома, мімівірус мае приблизно 911 генів, що кодують білок, що значно більше 4 необхідних кожному вірусу генів. Аналіз генома показав наявність генів, не присутніх ні у яких інших вірусів, зокрема, генів, що кодують аміноацил - тРНК - синтазу, і інших, виявлених тільки у організмів з клітинною будовою. Як і інші великі ДНК віруси, мімівірус містить набір генів для кодування ферментів вуглеводного, ліпідного і амінокислотного метаболізму, однак серед них є і такі гени, які не знайдені у інших вірусів.
Приблизно 10 % генома приходиться на некодуючі ділянки ДНК. Гіпотетично представники групи великих ядерно-цитоплазматичних вірусів (поксвіруси, ірідовіруси, фікоднавіруси) ведуть свій початок від ДНК-вмісних складніших (можливо клітинних) форм, таких як сучасні мікоплазми і рикетсії. На користь цієї гіпотези говорить наявність в геномі великих ДНК вірусів великої кількості "надлишкових" генів, що не є необхідними для розмноження і функціонально дублюючих господарські. Вважається, що ці гени є свого роду "реліктом", що залишився від періоду "самостійності".
Багато з вірусних генів не мають гомологів в геномах клітинних організмів, що вказує на еволюційну відособленість їх гіпотетичного предка від архей, прокаріот і еукаріот, що теоретично дозволяє виділити їх в четвертий домен живого.
Стадії реплікації мімівіруса все ще слабо вивчені. Відомо, що мімівірус приєднується до рецепторів на поверхні клітин амеби і потрапляє всередину клітини. Усередині вірус розпадається, а інфікована клітина продовжує нормальну життєдіяльність. Через приблизно 4 години в декількох місцях усередині амеби починають з'являтися скупчення речовини, через 8 годин після інфекції в клітині вже добре помітна безліч віріонів мимивируса. Цитоплазма продовжує наповнюватися новими віріонами і через 24 години після інфікування клітина розривається і вивільняє їх.
За допомогою електронної мікроскопії інфікованих клітин була виявлена агрегація капсиду мімівіруса і ядра клітини, встановлено утворення внутрішнього ліпідного шару вірусу з брунькуванням від ядра, а також знайдені частки, схожі з "вірусними фабриками", які утворюються при інфекціях іншими вірусами тієї ж групи.
Мімівірус та визначення поняття «життя»
Мімівірус має багато властивостей, які поміщають його на кордон живого і неживого. По своїх розмірах він перевершує деяких бактерій, таких як Rickettsia conorii або Tropheryma whipplei, містить геном, схожий за розміром з геномом багатьох бактерій (у тому числі вищеназваних), і має гени, не знайдені у інших вірусів, у тому числі кодуючі ферменти синтезу нуклеотидів і амінокислот, які відсутні навіть у деяких невеликих бактерій-внутрішньоклітинних паразитів. Це означає незалежність мімівіруса (на відміну від вказаних бактерій) від генома клітини-хазяїна, що кодує основні метаболічні шляхи. Проте мімівірус не має генів синтезу рибосомальных білків, через що він має необхідність в рибосомах хазяїна. Поєднання цих властивостей викликало в науковому середовищі спори, чи є мімівірус особливою формою життя, доменом, разом з еукаріотами, бактеріями і археями.
Проте, мимивирус не має гомеостазу, не відповідає на подразники, не росте і не розмножується самостійно (замість цього синтезується клітиною і самозбираєтся в ній з окремих компонентів), що типово для вірусів.
Гени, властиві мімівірусу (у тому числі кодуючі білки капсиду), зберігаються в безлічі вірусів, що вражають організми усіх трьох доменів. На підставі цього факту робиться припущення, що мімівірус пов'язаний з ДНК вірусами, які з'явилися одночасно з найбільш стародавніми організмами, що мають клітинну будову, і займають ключове місце в походженні життя на Землі.
Дослідження мімівірусів далеко не завершені. Останнім часом французькими дослідниками отримані дані про те, що і у вірусів можуть існувати свої "паразити". У мімівіруса виявлені "вірофаги", що дістали назву "супутники", размноження яких в клітинах амеби відбувається тільки у присутності мімівіруса і при цьому змінюється структура оболонки мімівіруса.
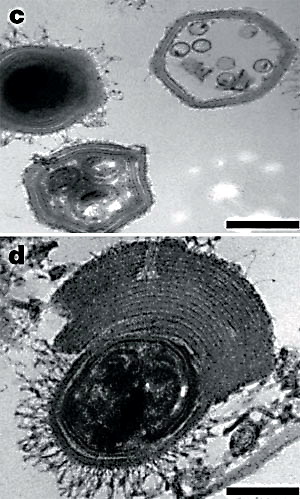
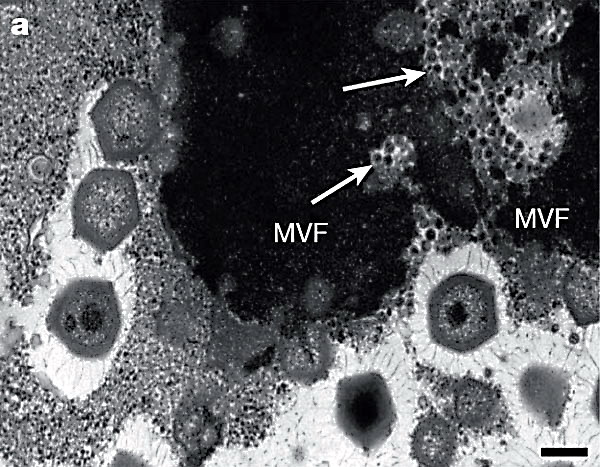
Рис. 27. Чорна область на знімку - це "вірусна фабрика" мімівіруса в цитоплазмі амеби. Великі шестикутні тіла - віріони мімівіруса на різних стадіях зрілости. Стрілками показані скупчення вірусів-супутників.
У "хворих" мімівірусів, уражених "супутниковою" інфекцією, часто спостерігається логічне потовщення оболонки. Довжина масштабної лінійки 200 нм.
Геном супутника є кільцевою молекулою ДНК. Вона складається з 18 343 пар нуклеотидів і містить 21 білок-кодуючий ген. Три з них, можливо, порівняно нещодавно були "запозичені" супутником у мімівіруса, тобто здобуті в результаті горизонтального обміну генами - це явище дуже широко поширене у світі вірусів. Ще один ген походить від вірусів - паразитів архебактерій, два інших - від поки що не описаної родини морських вірусів, про існування якої стало відомо в результаті широкомасштабних досліджень вірусної ДНК, витягнутої з морської води. Таким чином, супутник має хімерну природу; він "зібрав" свій геном, як мінімум, з трьох різних джерел.
Подібно до того, як бактеріофаги активно переносять гени від одних мікробів до інших, вірофаги можуть грати помітну роль в горизонтальному генетичному обміні між різними групами вірусів.
Висловлена точка зору може знайти своє підтвердження лише за наявності доказів вбудовування генома "вірофагів" у вірусні геноми, подібно до того, що можна спостерігати при явищі трансдукції - захоплення і перенесення бактеріофагами частини генома клітини - хазяїна.