Свойства плазмид.
Плазмиды-1)кольцевые 2-цепочечные мол-лы ДНК; 2)внехромосомный ген. элемент бактерий, способный к автономному сущ. и репликации, т.е. могут реплицироваться независимо от клеточной хромосомы.
Есть в клетках почти всех бакт., но в опред. усл. бакт. может жить без плазмид. В клетке есть высококопийные (10-100 копий) и низкокопийные(1-4 копии) плазмиды. У Е. Coli обычно 20 мелких копий, 1 или 2 большие. Размер - от 2х до сотен тыс. пар оснований. Доляплазмидной ДНК в клетке бакт. = 0,1-5 %всей ДНК клетки.
Плазмиды им. группы совместимости др. с др.: к одной группе несовместимости принадлежат плазмиды, кот.не могут сосущ. в 1й клетке; к разным группам –норм. сосущ. в 1й клетке независимо от числа копий.
Все плазмиды содержат сайт начала репликации (origin) в клетке хозяина. Одни плазмиды несут специфичный сайт инициации репликации и могут реплиц. только в клетках 1го вида(с узким спектром хозяев).У других этот сайт менее специфичен, и они реплиц. в разных бакт. клетках (с широким спектром хозяев).
Плазмиды содержат гены: инактивирование а/б; метаболизм прир. соед.; образ.токсинов. => группы плазмид по ф-ям: 1) F-плазмиды содержат инфу, обеспеч. их собственный перенос от клетки к клетке; 2) R-плазмиды несут гены устойчивости к а/б, т.е. распада/окисления а/б; 3) Плазмиды деградации несут гены утилизации необычных метаболитов; 4) Криптические – функциональные гены не обнаружены.
Природныеплазмиды часто бывают лишены обязат. для вектора свойств: небольшого размера,наличия уникального сайта рестрикции для вставки и селективных ген. маркеров для идентификации.
Роль ars последовательностей.
Посл-ти ДНК дрожжей, придающие плазмидамспос-ть реплицироваться в дрожжах, наз-сяARS – автономно реплицирующиесяпосл-ти. Встречаются в геноме дрожжей с той же частотой, что для точек начала репликации (1/40 тыс пар нукл., посчитано с пом. изотопа трития).
Для функц.ARSнеобх.посл-ть из 11 пар нуклеотидов: 5`-(A/T)TTTAT(A/G)TTT(A/T)-3`.
Искусственно сконструир. тандем из таких посл-тей при встраивании в плазмиду функционирует в дрожжевой клетке как ARS =>инициаторный белок (DNA-B), чтобы начать репликацию, должен узнать на ДНК располож. рядом др. с др. антипараллельные копии посл-ти 5 `…3 `, 3 `…5 `, т.е. ARSпосл-ти выступают в роли точек начала репликации, и содерж. их плазмиды способны самостоятельно реплицироваться в хозяйской клетке, не включаясь в состав хромосомы.
Метод выявления ARS элементов у дрожжей S.Cerevisiae:
В 1 из плазмидных векторов с геном аминок-ты гистидин вводят случайный фрагмент дрожжевой ДНК, в другой - ARS–фрагмент дрожжевой ДНК.Плазмиды вводят в дрожж. клетки, кот.не содержат ген гистидина. Дрожжи высаживают в чашки Петри с селективной средой без гистидина => в 1 случае вырастают редкие трансформанты сплазмидной ДНК, встроившиеся в хромосому дрожжей; во 2м - много трансформантов, содерж. кольцевую плазмидную ДНК, способную реплицироваться независимо от хозяйской хромосомы.
Получение к-днк методом пцр.
С пом. ПЦР можно получать комплементарные ДНК (кДНК), отвечающие 3'- или 5'- концевым участкам мРНК. Метод наз-ся RACE (от англ. rapidamplificationof с DNA ends) - быстрая амплификация (увеличение кол-ва) концов кДНКдля встраивания в вектор, а далее в составе вектора в клетку хозяина. Для провед. ПЦР-амплификации нужно знать нуклеотидную посл-ть кодирующей области мРНК- мишени, чтобы синтез-тьгенспецифичныйпраймер (GSP). Различают 3' RACE и 5' RACE методы.
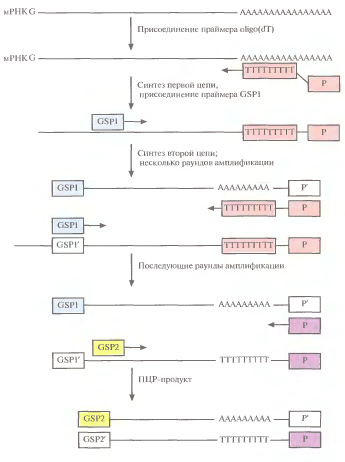
3' RACE:праймеромдля с-за первой цепи кДНК служат короткие олигонуклеотидные цепочки из остатков тимидина (около 15 звеньев) - oligo(dT), с присоед-м к нему вторым праймером (Р) (рис.1). Oligo(dT) спаривается с poly(A)- хвостом мРНК, и обратная транскриптаза синтез-ет цепь, комплементарную мРНК. Вторая цепь кДНК синтез.на первой при участии GSP, комплементарного кодирующей области данной мРНК, с помощью полимеразыTaq. По заверш. неск. раундов ПЦР, в которых исп-сь эти праймеры, добавляют вторую парупраймеров, которые связ. по соседству с первыми. Такие тесно расположенные праймеры называются внутренними. Вторая пара праймеровнеобх., т.к.без них нельзя амплифицировать полноразмерную молекулу-мишень. Конечным ПЦР-продуктом является кДНК, соотв. 3'-концу искомой мРНК.
5'RACE:праймером для с-за первой цепи кДНК служит GSP (рис. 2). Новую цепь обраб.концевойдезоксинуклеотидилтрансферазой в присутствии dATP (дезоксирибоаденинтрифосфата). Этот фермент присоед.дезосирибонуклеотиды к 3'-концу цепи. Т.к. в реакционной смеси есть только dATP, на этом конце появляется цепочка адениновых остатков — роlу(А)-хвост. С ним спаривается праймер P-oligo(dT), инициирующий синтез второй цепи. Проводят ограниченное число раундов ПЦР с указаннымипраймерами, а затем добавляют вторые праймеры и амплифицируюткДНК, отвечающую 5'-концу мРНК.
RACE-метод широко применяется по ряду причин. Обычно трудно обнаружить кДНК, соотв. мРНК, которой мало в данной ткани. С помощью RACE-метода можно быстро получить кДНК, отвеч. концевым участкам этой мРНК, и при необх. исп-ть их в качестве зондов для скрининга (отбора) кДНК- и геномных библиотек.
Рис.
1 (слева). 3 'RACE.
Первая цепь кДНКобраз.в рез-те обратной
транскрипции мРНК при участии
oligo(dT) в качестве праймера. Вторая цепь
синтез-ся на первой цепи как на матрице
с пом. полимеразы Taq и генспецифичногопраймера
(GSP1). В след.раундах ПЦР исп-сяпраймеры
GSP2 и Р.
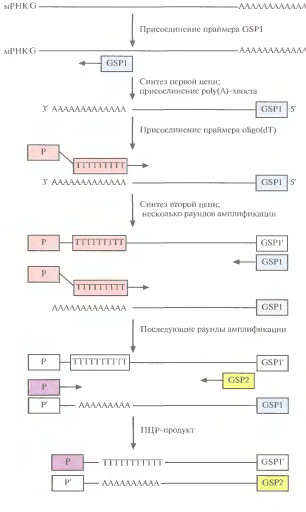
Рис. 2 (справа). ПЦР-амплификация кДНК, комплем. 5' концевой части мРНК первой цепи, осущ. обратной транскриптазой, инициируется праймером GSP. Затем к этой цепи с пом. концевой дезоксинуклеотидилтрансферазыприсоед-сяроlу(А)-хвост. Для синтеза второй цепи в качестве праймера используется oligo(dT). После неск. раундов амплификации добавляют вторые праймеры (GSP2 и Р)и получают кДНК,отвечающую 5'-концумРНК