

17.6. Гормоны поджелудочной железы
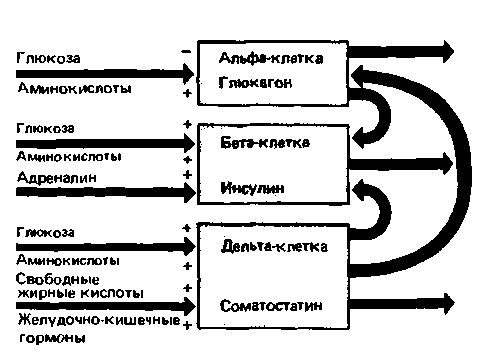
Гормоны, участвующие в поддержании постоянного уровня сахара в крови,- инсулин, глюкагон и соматостатин - секретируются островками Лангерганса в поджелудочной железе. Эти структуры представляют собой скопления эндокринных клеток, так называемых альфа-, бета- и дельта-клеток, разбросанных в виде групп по несколько тысяч клеток среди экзокринной ткани, составляющей основную массу поджелудочной железы. Около 60% островковых клеток приходится на бета-клетки, вырабатывающие инсулин, 25% - на альфа-клетки, секретирующие глюкагон, и остальные 15% - на дельта-клетки, выделяющие соматостатин. Регуляторные эффекты, определяющие секрецию перечисленных гормонов и зависящие от питательных веществ и паракринных влияний, схематически изображены на рис. 17.19. Для нормального функционирования островковых клеток необходим нормальный уровень тиреоидных и половых гормонов, а также кортикостероидов.
|
Рис. 17.19. Регуляция активности островковых клеток. Продуктами секреции клеток островков Лангерганса являются глюкагон, инсулин и соматостатин, секретируемые альфа-, бета- и дельта-клетками соответственно. Секреция инсулина стимулируется высокой концентрацией в крови глюкозы и аминокислот и угнетается адреналином. Секреторная активность альфа-клеток, вырабатывающих глюкагон, подавляется высокой концентрацией глюкозы в крови и стимулируется ее падением; аминокислоты оказывают противоположное действие. Секретируемый альфа-клетками глюкагон, действуя паракринным путем, угнетает секрецию инсулина бета-клеткам и. Секреция соматостатина дельта-клетками стимулируется высоким уровнем глюкозы, аминокислот и жирных кислот, а также гормонами желудочно-кишечного тракта. Паракринным путем соматостатин угнетает секрецию инсулина бета-клетками |
406 ЧАСТЬ IV. ПРОЦЕССЫ НЕРВНОЙ И ГУМОРАЛЬНОЙ РЕГУЛЯЦИИ
Инсулин
Химическое строение и действие на уровень сахара в крови. Инсулин, образующийся в бета-клетках поджелудочной железы, представляет собой полипептид, состоящий из двух пептидных цепей. А-цепъ, состоящая из 21 аминокислотного остатка, соединена дисульфидным мостиком с B-цепью, состоящей из 30 аминокислотных остатков. Инсулины животных разных видов лишь слегка различаются по аминокислотной последовательности и обладают одинаковым биологическим действием.
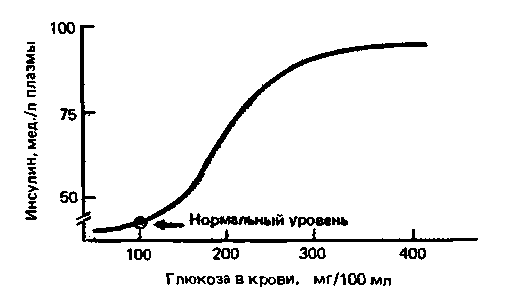
Суммарный результат различных метаболических эффектов инсулина сводится к понижению концентрации глюкозы в крови, составляющей в норме 0,8-1,0 г/л. При повышении уровня сахара в крови после приема углеводов происходит выброс инсулина. Под влиянием инсулина увеличивается поглощение глюкозы почти всеми клетками тела, и ее концентрация в крови опять снижается (рис. 17.20).
Действие на обмен глюкозы в печени. Печень это один из наиболее важных органов тела, запасающих глюкозу. Глюкоза может свободно диффундировать в клетки печени и выходить из них, когда ее содержание в крови снижается. В клетках печени глюкоза под влиянием инсулина превращается в гликоген, и ее содержание в крови снижается. Один из главных биохимических эффектов инсулина состоит в активации фермента глюкокиназы, катализирующей фосфорилирование глюкозы, которая поступает в клетки печени. Инсулин активирует и другие ферменты, в том числе фосфофруктокиназу и гликогенсинтетазу, катализирующую полимеризацию фосфорилированной глюкозы с образованием гликогена. Наконец, инсулин ингибирует ферменты, расщепляющие гликоген (фосфорилазы), бла-
|
Рис. 17.20. Зависимость концентрации инсулина в крови от содержания в ней глюкозы. При нормальной концентрации глюкозы (около 100 мг/100 мл) инсулин в плазме практически не определяется, но при повышении уровня сахара в крови в физиологических условиях концентрация инсулина резко возрастает |
годаря чему высокий уровень инсулина способствует консервации гликогена. За счет быстрого синтеза гликогена и подавления гликогенолиза концентрация глюкозы в крови, повышающаяся после приема пищи, быстро возвращается к нормальному уровню. В результате удаляется основной стимул секреции инсулина, и содержание последнего в крови также нормализуется.
Когда организму требуется энергия в промежутках между приемами пищи, гликоген опять превращается в глюкозу. Концентрация инсулина в крови в этот период очень мала, поэтому фосфорилаза находится в активном состоянии и превращает гликоген в глюкозофосфат, который дефосфорилируется глюкозофосфатазой. Образующаяся при этом глюкоза может свободно выходить из клетки путем диффузии. Таким образом поддерживается постоянный уровень глюкозы в крови между приемами пищи. При нормальном питании около 60% глюкозы, потребляемой человеком с пищей, временно запасается в печени, с тем чтобы затем быстро высвобождаться за счет расщепления гликогена [3, 5, 6, 11].
Действие на обмен глюкозы в мышечных клетках.
При низком содержании инсулина в крови мышечные клетки в норме непроницаемы для глюкозы и всю необходимую энергию получают за счет окисления жирных кислот. Увеличение концентрации инсулина, вызванное повышением уровня глюкозы в крови после приема пищи, делает мышечные клетки проницаемыми для глюкозы, которая используется затем в качестве источника энергии. Однако при очень высокой мышечной активности мембраны клеток становятся проницаемыми для глюкозы и в отсутствие инсулина. В этом случае потребность работающей мышцы в глюкозе как энергетическом субстрате удовлетворяется даже при базальном уровне инсулина. Детально этот механизм еще не изучен. Когда мышца находится в неактивном состоянии, в ней сразу после приема пищи, т. е. при высоких концентрациях инсулина и глюкозы, тоже образуется и сохраняется небольшое количество гликогена. При острой необходимости этот гликоген опять превращается в глюкозу, которая используется мышечными клетками (с. 84 и 698). Как правило, глюкоза не выделяется обратно в кровь и не играет никакой роли в регуляции уровня сахара в крови после приема пищи.
Связываясь с рецептором на поверхности мышечных клеток, инсулин активирует механизм транспорта глюкозы через мембрану. Пока неизвестно, служит ли белком-переносчиком сам инсулин-рецепторный комплекс или он активирует другую транспортную систему.