Глава 17. Эндокринология 397
Патофизиологические аспекты. Результаты опытов на животных позволяют предположить, что выделяемый гипоталамусом дофамин угнетает также клетки, секретирующие ЛГ-РГ, поскольку при активации дофаминергических клеток в гипоталамусе секреция ЛГ-РГ уменьшается. Соответственно и секреция ЛГ и ФСГ гипофизом падает ниже нормы (очевидно, за счет угнетения эпизодической секреции; с. 831), что приводит к нарушению менструального цикла. Такое состояние называется лактогенной аменореей. Достаточно хорошо этот механизм работает только у часто кормящих матерей: если младенец питается не только грудным молоком, то этот механизм не обеспечивает матери контрацептивной защиты.
Серьезным видом патологии является гиперпролактинемическая аменорея. Сравнительно часто у женщин спонтанно развивается доброкачественная аденома гипофиза, клетки которой секретируют большие количества пролактина. Как правило, эти клетки сохраняют чувствительность к дофамину, но, по-видимому, не находятся под гипоталамическим контролем, осуществляемым через воротные вены, поскольку снабжение их кровью происходит целиком через гипофизарные артерии. Высокая концентрация пролактина в крови нередко вызывает лактацию, которая у некормящих женщин называется галактореей. Посредством механизма обратной связи пролактин стимулирует дофаминергические нейроны гипоталамуса, и в норме этот эффект противодействует избыточной секреции пролактина. Но тот дофамин, который поступает из гипоталамуса в гипофиз, не достигает опухолевых клеток, поэтому их активность не подавляется. В то же время избыток дофамина угнетает дофаминергические нейроны, секретирующие ЛГ-РГ, что приводит к подавлению секреции ЛГ-РГ, нарушению циклической активности и развитию аменореи. Это нарушение легко снимается введением агонистов дофамина, которые через общую систему кровообращения могут достигать опухолевых клеток гипофиза, секретирующих пролактин, и подавлять их секреторную активность. После нормализации секреции пролактина всегда нормализуется и половой цикл.
Тоническое угнетение дофамином секреции пролактина имеет еще один важный фармакологический аспект. Многие лекарства (очень часто психотропного действия) блокируют рецепторы дофамина, образуя с ними неактивный комплекс, неспособный запускать те внутриклеточные механизмы, которые активируются комплексом рецептора с самим дофамином. Такие вещества называют антагонистами дофамина, поскольку они препятствуют его действию, конкурируя с ним за рецепторы. Именно такой эффект и требуется от этих препаратов, но одновременно он приводит и к повышению секреции пролактииа, следствием чего могут быть галакторея и аменорея.
17.4. Система щитовидной железы Образование и секреция тиреоидных гормонов
Регуляция синтеза гормонов. В 1969 г. две группы исследователей, руководимые Шалли [38] и Гьюллемином [20], одновременно и независимо одна от другой выделили из гипоталамуса соответственно
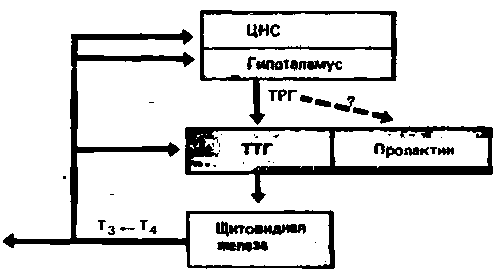
свиней и овец трипептид, стимулирующий секрецию ТТГ гипофизом. Это был первый рилизинг-гормон, изученный химически и полученный синтетическим путем. Данный пептид образуется в нейронах гипоталамуса и высвобождается в воротную систему (рис. 17.13). Достигнув гипофизарных клеток, секретирующих ТТГ, этот тиреотрорин-рилизинг-гормон (ТТГ-Ρ Г) стимулирует его секрецию. В свою очередь ТТГ действует на щитовидную железу и стимулирует синтез и секрецию гормонов тироксина и трийодтиронина. По механизму отрицательной обратной связи тиреоидные гормоны влияют на гипоталамус и гипофиз, и, когда их концентрация в крови бывает максимальной, секреция ТТГ снижается до минимума. Напротив, низкая концентрация тиреоидных гормонов в крови обусловливает высокую скорость секреции ТТГ. Тот факт, что у животных с нарушенной в результате повреждения гипоталамуса секрецией ТТГ-РГ сохраняется достаточно хорошая регуляция уровня тиреоидных гормонов в крови, доказывает, что механизм отрицательной обратной связи действует главным образом на уровне гипофиза. Синтетический ТТГ-РГ может быть использован для диагностических целей, поскольку при введении он вызывает быстрое увеличение секреции ТТГ гипофизом. Удивительно, однако, что при этом он стимулирует также секрецию пролактина (с. 396).
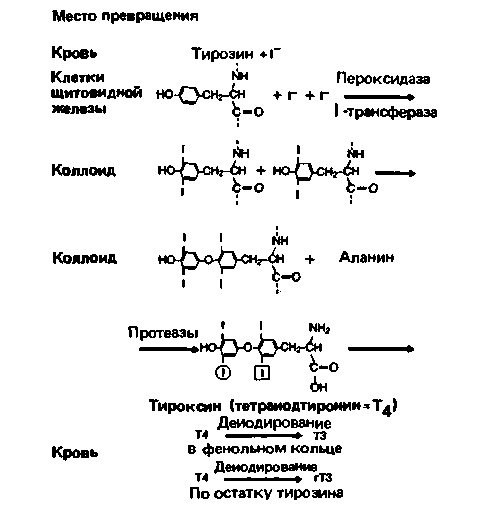
Синтез и секреция тиреоидных гормонов. Чтобы происходил синтез тиреоидных гормонов, суточное потребление иода должно составлять около 150 мкг. Поступающий с пищей иод всасывается в кишечнике в кровь и поглощается из нее клетками щитовидной
|
Рис. 17.13. Система гипоталамус-гипофиз-щитовидная железа. Нейроны гипоталамуса секретируют тиреотропин-рилизинг-гормон (ТРГ), который по воротной системе поступает в гипофиз, и стимулирует секрецию тиреотропного гормона (ТТГ). ТТГ индуцирует в щитовидной железе образование тироксина (Т4) и трииодтиронина (Т3). Действуя по механизму отрицательной обратной связи на гипофиз и, возможно, на гипоталамус, Т3 угнетает секрецию ТТГ. Пока неясно, может ли в физиологических условиях высвобождение гипоталамусом ТРГ стимулировать также секрецию пролактина |
jpg часть iv. процессы нервной и гуморальной регуляции
|
Рис. 17.14. Образование, хранение и высвобождение тиреоидных гормонов. Большая часть суточной потребности в иоде покрывается за счет всасывания его в кишечнике в виде ионов. Последние поглощаются из крови клетками щитовидной железы и окисляются пероксидазой до молекулярного иода. В щитовидной железе образуется тиреоглобулин, богатый остатками тирозина, с которыми и взаимодействуют молекулы иода. В результате конденсации двух молекул дииодтирозина, находящихся в составе тиреоглобулина, образуется тироксин, который остается связанным с тиреоглобулином и в таком виде поглощается из коллоида клетками щитовидной железы. В этих клетках тироксин отщепляется под действием ферментов от тиреоглобулина и выделяется в кровоток. На периферии тироксин подвергается деиодированию с образованием трииодтиронина активного тиреоидного гормона; в самой щитовидной железе образуется лишь очень небольшая часть трииодтиронина. Некоторое количество иода, освобождающегося при деиодировании тироксина, поступает обратно в щитовидную железу, но большая часть его выводится почками |
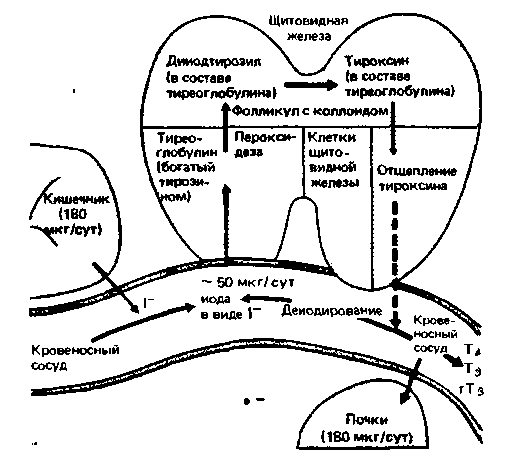
железы (рис. 17.14). Все описанные ниже процессы стимулируются ТТГ. Прежде всего следует отметить, что концентрация иода в клетках выше, чем в крови, поэтому он не может поступать в них путем диффузии, а должен активно переноситься с затратой энергии. В клетках щитовидной железы синтезируется высокомолекулярный белок - тиреоглобулин, содержащий множество остатков тирозина. Поступивший в клетку иод включается в 3-е и/или 5-е положения бензольного кольца тирозина, входящего в состав тиреоглобулина, с образованием моно- или дииодтирозила (рис. 17.14). В условиях in vitro иодирование может идти спонтанно, но в щитовидной железе иодид окисляется ферментом пероксидазой до активного атомарного иода, который
и соединяется с тирозильными радикалами тиреоглобулина. Иодированные тирозильные радикалы конденсируются с образованием тироксина (тетраиодтиронина, Т4); при этом образуются также и небольшие количества трииодтиронина (Т3). В конечном счете Т3 и Т4 высвобождаются из клетки в составе молекулы тиреоглобулина и попадают в фолликулы щитовидной железы, представляющие собой пузырьки, образованные секреторными клетками. В фолликуле тиреоидные гормоны хранятся в составе тиреоглобулина, образующего гелеобразное вещество, называемое коллоидом.
Для выделения тиреоидных гормонов коллоид должен быть захвачен клетками железы, что осуществляется путем пиноцитоза. В клетках щитовидной железы тиреоглобулин расщепляется с образованием свободных молекул тироксина и трииодтиронина, которые диффундируют в кровь. В крови они связываются нековалентно с белками плазмы, и лишь небольшая часть их присутствует в крови в свободном виде. Среди белков плазмы, связывающих тиреоидные гормоны, наиболее важную роль играют тироксин-связывающий глобулин (ТСГ), тироксин-связывающий преальбумин и ряд других альбуминов.
Весь тироксин, присутствующий в крови, секре-
|
Рис. 17.15. Структурные формулы тироксина (Т4), трииодтиронина (Т3) и ревертированного трииодтиронина (NTj). Биологической активностью обладает только Т3. Щитовидная железа высвобождает значительно больше Т4 по сравнению с Т3, но Т4 превращается на периферии в Т3 в результате деиодирования по фенольному кольцу. При деиодировании в бензольном кольце образуется гТ3, не обладающий активностью |