

Глава 22. Транспорт газов крови 619
ви обусловлены способностью аминокислот ионизироваться. Концевые карбокси- и аминогруппы белковых цепей играют в этом отношении незначительную роль, так как таких групп мало и их рК' существенно отличаются от pH крови. Значительно больший вклад в создание буферной емкости белковой системы вносят боковые группы, способные ионизироваться, и особенно имидазольное кольцо гистидина. К буферным белкам относятся как белки плазмы (в частности, альбумин), так и содержащийся в эритроцитах гемоглобин. На долю последнего приходится большая часть буферной емкости белковой системы, что связано как со значительной концентрацией гемоглобина, так и с относительно высоким содержанием в нем гистидина.
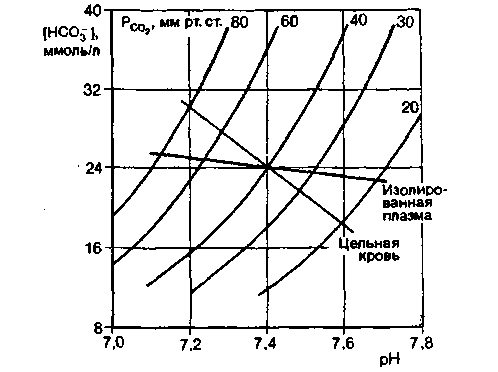
Особенно наглядно роль гемоглобина в создании буферной емкости крови можно показать, если сравнить сдвиги pH в ллазме и цельной крови, вызываемые одинаковыми изменениями напряжения СО2. Результаты такого сравнения удобно представить в виде графика зависимости pH от [HCO3− ], на который нанесены кривые для постоянного напряжения СО2, рассчитанные в соответствии с уравнением Гендерсона-Гассельбальха [уравнение (19)]. Измерив при разных значениях напряжения СО2 pH и содержание HCO3− в плазме и в цельной крови, где происходит обмен ионов между плазмой и эритроцитами, строят так называемые равновесные кривые СО2 для изолированной плазмы и цельной крови (рис. 22.12). Чем круче наклон кривой для цельной крови, тем больший вклад вносит в буферную емкость крови гемоглобин, и чем больше угол наклона кривой буферной емкости, тем меньше изменения pH при данном увеличении или уменьшении напряжения СО2 .
|
Рис. 22.12. Равновесные кривые СО2 для изолированной плазмы и цельной крови. По оси ординат -содержание в плазме бикарбоната, по оси абсцисс pH. Приведены также кривые для постоянного напряжения СО2. Поскольку в цельной крови между плазмой и эритроцитами происходит обмен ионами, буферная емкость плазмы добавляется к буферной емкости гемоглобина и наклон соответствующей равновесной кривой становится круче |
|
Рис. 22.13. Буферные кривые для недиффундирующих буферных систем эритроцита на основе гемоглобина (при незначительном участии АТР и 2,3-ДФГ) (по [19] с изменениями). По оси ординат-отношение содержания недиффундирующих буферных анионов Pi к содержанию гемоглобина; по оси абсцисс-pH внутри эритроцита. ИHbO2 и ИHb изоэлектрические точки. Благодаря тому что буферная кривая для Hb смещена вправо относительно буферной кривой для НbО2. при полной деэоксигенации гемоглобина 1 ммоль Hb может дополнительно связывать 0,45 ммоль Н+ без изменения ρΗ внутри эритроцита |
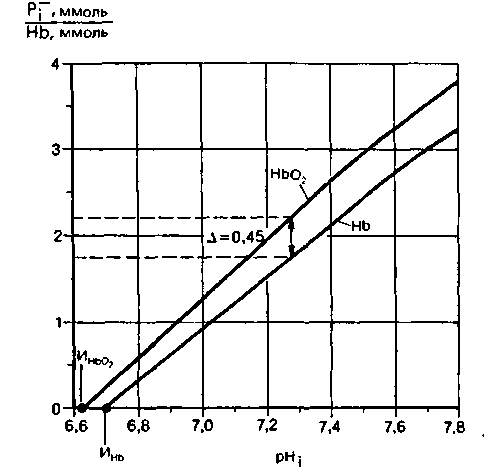
Гемоглобин играет также важную роль в буферной емкости крови благодаря изменению его кислотных свойств при оксигенации и дезоксигенации. Эта зависимость выражена на рис. 22.13, где приведены буферные кривые (кривые титрования) для недиффундирующих буферных систем эритроцита на основе гемоглобина в оксигенированном и дезоксигенированном состояниях. Видно, что в физиологическом диапазоне pH оксигемоглобин является более сильной кислотой, чем дезоксигемоглобин. Такая разница обусловлена главным образом влиянием кислорода, связанного с железом, на связывание Н + соседними имидазольными группами гистидина. Благодаря этому влиянию гемоглобин, освобождая в тканях кислород, приобретает большую способность к связыванию ионов Η+, образующихся при одновременном поглощении СО2. При поглощении кислорода кровью в легких происходят обратные процессы. Таким образом, обмен О2 усиливает буферный эффект гемоглобина.
Буферные основания. Буферные свойства крови обусловлены суммарным эффектом всех анионных
620 ЧАСТЬ VI. ДЫХАНИЕ
групп слабых кислот; важнейшими из них являются бикарбонат и анионные группы белков («протеинаты»). Все эти анионы, обладающие буферными эффектами, называют буферными основаниями (по международной номенклатуре-buffer bases, BB) [10].
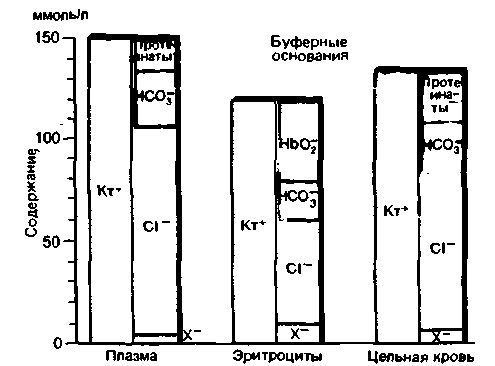
На рис. 22.14 приведены концентрации различных ионов в плазме, эритроцитах и цельной крови человека (для артериальной крови). Высота столбиков пропорциональна концентрации ионов. Верхняя часть столбиков, относящихся к анионам, соответствует буферным основаниям (темно-красный цвет), а нижняя часть-всем анионам сильных кислот (кроме Сl−), не обладающих буферными свойствами, таким как SO42− и органические анионы (обозначенные вместе Х~). Из рисунка видно, что в плазме основную часть буферных оснований составляют ионы НСО3− , а в эритроцитах-протеинаты. В цельной крови более чем 1/3 всех анионов обладают буферным действием.
Концентрация буферных оснований в артериальной крови составляет примерно 48 ммоль/л. Очень важно, что эта величина не изменяется при сдвигах напряжения СО2. Рассмотрим причины такого постоянства. Предположим, например, что напряжение СО2 возрастает. При этом образуются равные количества Н+ и HCO3−. Ионы Н+ почти полностью связываются протеинатами, переходящими при этом в недиссоциированную форму. В результате концентрация протеинатов снижается настолько, насколько увеличивается концентрация бикарбоната (рис. 22.15).
|
Рис. 22.14. Содержание ионов в плазме, эритроцитах и цельной крови. К буферным основаниям относятся анионы буферных систем (темно-красный столбик); фосфаты не изображены, так как их содержание очень низко. Х− - анионы всех сильных кислот, не обладающих буферными свойствами, за исключением Cl−; Kt+ -суммарная концентрация катионов |
|
Рис. 22.15. Зависимость концентраций протеинатов и бикарбоната от РСO2 . Суммарное содержание двух буферных оснований остается постоянным (в норме48 ммоль/л) |
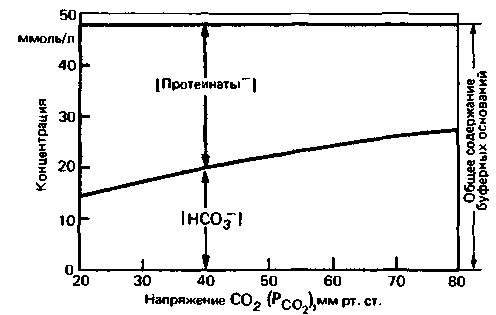
Подобные концентрационные сдвиги в буферных системах не взаимоуравновешиваются полностью, так как некоторые протоны остаются в свободной форме, изменяя тем самым pH. Так, если PCO2 возрастает от 40 до 50 мм рт. ст., то pH внутри эритроцитов снижается на 0,06, а в плазме- на 0,1. Это означает, что концентрация ионов Н+ увеличивается примерно на 10−5 ммоль/л. Однако этот сдвиг очень мал и не влияет на характер графика, приведенного на рис. 22.15.
Поскольку суммарная концентрация буферных оснований крови не зависит от РCO2 , по ней можно судить о сдвигах кислотно-щелочного равновесия, вызванных увеличением или уменьшением содержания нелетучих кислот в крови. Отклонение концентрации буферных оснований от нормального уровня (48 ммоль/л) называется избытком оснований (по международной номенклатуре-base excess, BE). Таким образом, в норме BE по определению равен нулю. При патологическом увеличении содержания буферных оснований BE становится положительным, а при снижении - отрицательным. В последнем случае можно использовать термин «дефицит оснований», так как выражение «отрицательный избыток оснований» с семантической точки зрения бессмысленно.