

Глава 21. Легочное дыхание 599
|
Рис. 21.32. Каротидные (А) и аортальные (Б) тельца и отходящие от них афферентные волокна. В. Зависимость частоты разрядов хеморецептора от напряжения 02 в артериальной крови. Приведена запись импульсации в синокаротидном нерве при перфузии изолированного каротидного синуса кошки кровью с различным напряжением 02 (РаО2 ) при постоянном напряжении С02 (33 мм рт. ст.) и pH 7,33. По оси ординат отложена суммарная импульсация в синокаротидном нерве в процентах от максимальной из возможных (f/fmax) [35] |
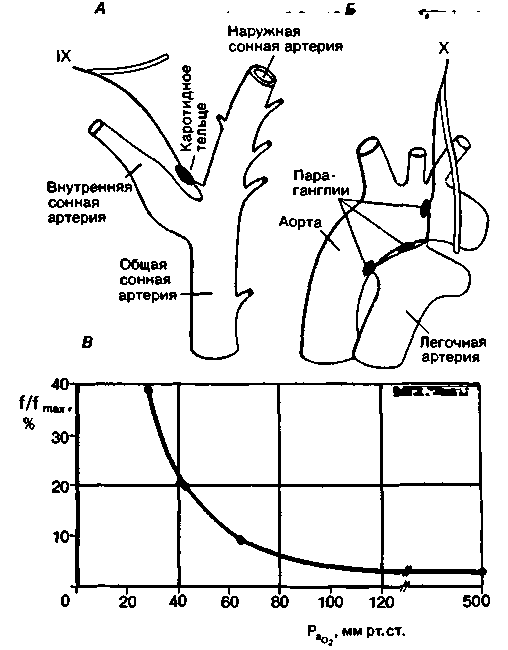
каротидное тельце иннервируется ветвью языкоглоточного нерва. Хеморецепторы обнаруживаются также в параганглиях дуги аорты (иногда их называют аортальными тельцами); импульсы от них поступают в ЦНС по афферентным волокнам блуждающего нерва. Параганглии кровоснабжаются за счет мелких коллатеральных артерий.
Хеморецепторы, расположенные во всех этих образованиях, возбуждаются (т.е. частота импульсации от них возрастает) при снижении напряжения О2, повышении напряжения СО2 или уменьшении pH. Это можно показать в опыте на животном, регистрируя изменения активности эфферентных нервных волокон в ответ на сдвиги показателей химического состава крови (рис. 21.32, В). В таких опытах артерии, снабжающие параганглии. перфузируются кровью, в которой установлены те или
иные уровни РО2 , РСО2 и pH. Роль хеморецепторов в осуществлении регуляторного влияния химических факторов можно изучать также путем денервации этих рецепторов или холодовой блокады их активности.
В подобных экспериментах было показано, что влияние О2 опосредовано исключительно периферическими хеморецепторами. При нормальном напряжении О2 в артериальной крови в волокнах этих рецепторов регистрируется некоторая фоновая импульсация: при снижении РО2 она повышается, а при его увеличении уменьшается. На активность рецепторов влияют также РСО2 и pH артериальной крови, однако роль этих сигналов в центральной регуляции дыхания относительно ограничена.
Чувствительность центральных структур к химическим факторам. Влияние СО2 и pH на дыхание опосредованно главным образом их действием на особые структуры ствола мозга, обладающие хемочувствительностью. РСО2 и pH артериальной крови оказывают неодинаковый эффект на дыхание (рис. 21.31). Из этого вовсе не следует, что в стволе мозга содержатся хеморецепторные образования двух типов; одни чувствительные к СО2, а другие-к ионам Н+ . Возможно, что все эти образования воспринимают только ионы Н+ , а действие СО2 связано с образованием этих ионов. В этом случае разница в эффектах РСО2 и pH артериальной крови может быть связана с различиями в сопротивлении транспорту СО2 и ионов Н+. Известно, что СО2 очень быстро диффундирует из крови в ткани мозга, тогда как ионы Н+ с трудом проникают через биологические мембраны. Данные многочисленных экспериментов свидетельствуют в пользу гипотезы о том, что центральные нервные структуры, участвующие в регуляции дыхания, чувствительны исключительно к ионам Н+ [38, 42].
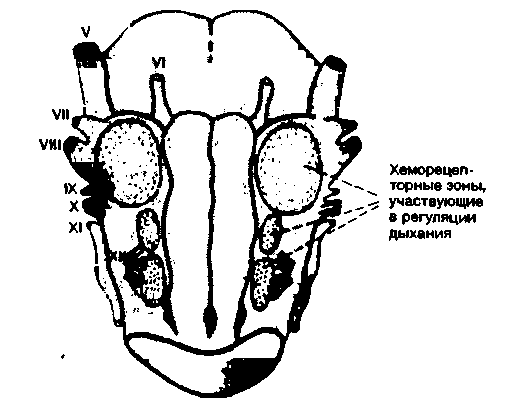
В настоящее время полагают, что основным химическим фактором, влияющим на дыхание, является содержание ионов Н+ в межклеточной жидкости ствола мозга. По-видимому, эта жидкость сходна со спинномозговой жидкостью (СМЖ), в связи с чем сдвиги в составе СМЖ также могут оказывать влияние на дыхание. Действительно, в опытах с изменением состава раствора, перфузирующего желудочки мозга, было обнаружено, что вентиляция легких зависит от pH этого раствора. Можно более точно определить локализацию участков, чувствительных к ионам Н+, на поверхности ствола мозга [38, 42]. Как показано на рис. 21.33, на вентральной поверхности продолговатого мозга обнаружены три таких участка: они расположены около корешков блуждающего и подъязычного нервов. Нанесение на эти участки кислот сопровождается увеличением вентиляции легких.
600 ЧАСТЬ VI. ДЫХАНИЕ
|
Рис. 21.33. Хеморецепторные зоны, участвующие в регуляции дыхания, на вентральной поверхности продолговатого мозга кошки [38, 42] |
Система химической регуляции дыхания. На рис. 21.34 схематично изображена вся регуляторная система, отвечающая за поддержание постоянства уровней дыхательных газов и pH в артериальной крови. Регулируемые переменные РО2 РСО2 и pH контролируются периферическими и центральными хемочувствительными структурами. Когда сенсоры регистрируют отклонение какой-либо из этих переменных от заданного значения (уставки, значения, заложенного в регуляторном контуре), они вызывают изменение активности нейронов продолговатого мозга. Это приводит к изменению сократительной
активности дыхательных мышц, в результате которого возникшее отклонение корригируется.
Эти три регулируемые переменные влияют на минутный объем дыхания по-разному. В норме главную роль играет парциальное давление СО2 в артериальной крови. Однако при утрате чувствительности центральных хеморецепторов (в результате хронических легочных заболеваний, сопровождающихся хронической гиперкапнией, или отравления барбитуратами и т. д.) основным стимулирующим фактором спонтанного дыхания становится артериальная гипоксия, возбуждающая центральный контроллер через периферические хеморецепторы. Если такому больному дать дышать чистым кислородом, то основной стимулятор дыхания устраняется, и больной может погибнуть в результате остановки дыхания. В этих случаях следует давать чистый кислород лишь при искусственном дыхании (или же при наличии «под рукой» аппарата искусственной вентиляции легких). При нарушениях кислотно-щелочного равновесия (с. 621) главной целью системы регуляции дыхания становится коррекция pH артериальной крови. Так, в ответ на метаболический ацидоз (снижение pH) возникает гипервентиляция; выделение СО2 увеличивается, и pH возвращается к нормальному значению.