

§ 3. Ассоциативные системы и сенсорная функция мозга
Прежде чем произойдет какая-нибудь приспособительная реакция, осуществляв ется процесс восприятия сигналов. Но живой организм заинтересован пе в самих воздействиях как таковых, а в том, о чем они сигнализируют, и соответственно не в оценке их физических параметров, а в тех соотношениях, которые с их помощью передаются. Вместе с тем попятно, что отнесение сигнала к тому или иному классу, то есть его распознавание, неизбежно основано на выделении ряда физических характеристик, образующих область признаков данного класса сигналов.
Из сказанного выше следует, что сенсорная функция мозга заключается в определении сигнальной (биологической) значимости сенсорных стимулов на основе анализа их физических характеристик. Под биологической значимостью понимается направленность реакций живого организма, которая определяется его доминирующей мотивацией, возникающей при отклонении параметров устойчивого неравновесия, а также информацией, извлекаемой из окружающем! среды » прошлого жизненного опыта. Для оценки биологической значимости сенсорны*--сигналов анализ их физических характеристик является необходимой, по недостаточной операцией. Биологическая значимость сигнала реализуется в актива-, ции некоей совокупности эффекторных аппаратов. Следовательно извлечении биологически полезной информации с целью ее использования для формирования двигательных актов основано на преобразовании входной кодовой комбинации активности нервных элементов сенсорных систем в реакцию исполнительных аппаратов, что по своей сути представляет процесс декодирования сенсорных сообщений. Тогда оценка биологической значимости сигнала сводится к установлению соответствующей закономерности взаимодействия нервных элементов сенсорных и двигательных систем мозга.
Выход на эффекторпые элементы должен обладать свойством избирательности, чтобы избавить систему от неадекватных реакций. Выделение экологически значимых признаков (обычно достаточно простых) у животных низких уровней эволюционного развития может осуществляться в самих сенсорных системах (иногда уже на рецепторпом уровне) вследствие жесткой организации нервных связей. Однако у более высокоорганизованных животных (хищных, приматов) эта операция уже пе может ограничиться специфическими образованиями сенсорных систем. Ведь по мере развития уровня организации животных рецентуар стимулов, имеющих сигнальное значение, как правило, расширяется. Поэтому возрастание активности организма как системы должно быть связано с возможностью перехода избирательного приема от одних сигналов к другим и в соответствии с этим — с появлением новых впутримозговьгх конструкций.
Нервные образования, входящие в систему оценки биологической значимости сигналов, должны удов;1етворить ряду требований. Во-первых, они должны быть связаны с различными сенсорными системами. Во-вторых, они должны быть связаны с мотивациогенпыми структурами лимбической системы, ибо значимость тех или иных сигналов определяется па основе доминирующей мотивации. В-третьих, учитывая, что значимость одного и того же сенсорного стимула зависят от всей окружающей ситуации, то есть от показаний других сенсорных систем, следует допустить Наличие конвергенции полимодалыюй имнульсации и способность к пластическим перестройкам активности нейронов. Наконец, в-четвертых, эта система должна быть связана с регуляцией целостных двигательных актов па основе избирательного характера реагирования. Изложенным требованиям удовлетворяют таламокортикальные ассоциативные системы мозга; таламопариетальпая и таламофроптальпая.
Получены доказательства (А. М. Ивапицкий, 1976) того, что ранние компоненты суммарного вызванного потенциала мозга человека отражают поступление в кору информации о физических характеристиках сенсорных сигналов, а поздние — являются отражением значимости сигнала для организма. У больных с психическими расстройствами, имеющих Определенный дефицит одного из видов информации, возникают стойкие изменения соответствующих фаз вызванных но-тенциаюв. В особенности выделяются ассоциативные области иеокортекса в связи с выраженной конвергенцией к ним разномодалыюй имнульсации как основы оценки сигнальной значимости стимула в контексте соответствующих воздействий.
Именно этот механизм у высших позвоночных способствует селективному отбору одних видов информации с одновременным сопряженным торможением Других сенсорных влияний. При этом у животных доминирующая мотивация возникает из биологической потребности в результате сдвигов гомеостаза. Однако Доминирующая мотивация формируется при воздействии стимулов, ранее связапных с определенными подкреплениями. Живая система способна предвидеть изменения гомеостаза с помощью сш налов, предвосхищающих сдвиг параметров от нормы или возвращение к ней. В связи с этим другие сенсорные стимулы способны изменять значимость сигнала, если они меняют доминирующую модернизацию. И тогда значимый стимул является сигналом, реагирование на который сопряжено с большей вероятностью достижения желаемого результата в соответствии с доминирующей мотивацией и прошлым жизненным опытом. Наличие сопряженных признаков в комплексном сенсорном воздействии позволяет уменьшить неопределенность выбора поведенческих реакций, необходимых для получения требуемого результата.
Роль ассоциативных систем в оценке биологической значимости сенсорных сигналов отчетливее видна по отношению к зрению и слуху. Конечно, сигнальное значение имеют все сенсорные стимулы, но способностью опережающего воздействия обладают в первую очередь зрительные, слуховые, а также обонятельные сигналы. Особенностью структурной организации телецентивных входов в ассоциативные системы является множественность нулей проведен ия к ним слуховых и зрительных афферентации.
Механизмы
конвергенции обеспечивают
полифункциональность одного и того
же сигнала, то есть зависимость его
смысла от ситуации, в которой он
возникает.
вании однообразной ритмической стимуляции Степень развития привыкания находится в прямой зависимости от биологической значимости применяемого сигнала. Например, стабильные ответы нейронов лобных долей у бодрствующих обезьян возникают лишь в том случае, если раздражители прямо или косвенно связаны с эмоционалыю-мотивациопной сферой животного, необычны для него или сигнализируют об опасности, то есть сопровождаются чет кой ориентировочной реакцией. Привыкание ответов наблюдается в тех случаях, когда используемые раздражители являются индифферентными для животного или хорошо знакомы ему (рис. 103). Если же эти агенты сопровождаются биологически значИ мым подкреплением по правилам выработки условного рефлекса, то они ириобре тают свойства условного сигнала с одновременной стабилизацией реакций неиро нов лобной коры в ответ на их применение.
Как привыкание, так и сенситизация отражают дистрибутивный характер Да пых пластических реакций мозга и совпадают с закономерностями проявления ориентировочного рефлекса. Поэтому предполагается, что «новизна» являе ^ тем информативным признаком сенсорных стимулов, с которого начинается
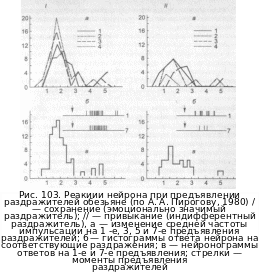
оценка
в ассоциативных системах мозга.
Привыкание к повторным стимулам,
вероятно, связано с процессами
формирования следов кратковременной
памяти (Е. Н. Соколов, 1981).
С помощью ориентировочной реакции устанавливаются соответствующие воспринимающие поверхности для наилучшего восприятия внешнего раздражения. При размещении источника телецентивных стимулов но средней линии головы наблюдается их наиболее высокое пространственное разрешение. Избирательность к этой физиологически значимой области пространства применительно к акустическим сигналам обнаружена для нейронов мозжечка и сепсомоторной ассоциативной коры (Я. А. Альтман, 1972; Г. А. Куликов, 1989). Участие фронтальной и теменной областей коры в обеспечении точности ориентировочных реакций Установлено и для зрительных сигналов, по отношению к которым отмечена и "Ространствеиная, и дирекциональная избирательная чувствительность соответствующих нейронов. Более того, после повреждения теменной ассоциативной об-Ласти отчетливо выступает нарушение пространственного восприятия зритель-"Ых стимулов и адекватных двигательных реакций, то есть нарушения прострап-Cl венного согласования зрительно-моторной координации.
Система, оценивающая биологическую значимость сенсорных стимулов, долж-1а °бладать избирательным характером реагирования па их отдельные значимые признаки и/или совокупность последних. Поэтому логичен вопрос о том, как отражаются в реакциях ассоциативных структур мозга различные физические параметры сенсорных сигналов.
Известно, что нейроны фронтальной ассоциативной коры принимают зрительную имнульсацию, пе связанную с поточечным описанием контура зрительных сигналов, но отражающую их временные и пространственные параметры. Кроме того, показано, что нейроны фронтальной коры обладают свойством избирательного реагирования па биологически значимые признаки коммуникативных акустических сигналов. Существенно то, что для млекопитающих (в отличие от низших позвоночных) лишь небольшое число сигналов может иметь четкое «ключевое» значение, которое в большей мере определяется доминирующим мотива-циопным фоном для данного животного. Поэтому становится понятным участие в оценке биологической значимости сигнала таламофропталыюй ассоциативной системы, тесно связанной с лимбическими структурами мозга, имеющими прямое отношение к формированию мотивациопиых состояний (Г. А. Куликов, 1989).
В связи с этим закономерен вопрос, как отражается в характере электрических реакций фронтальной коры процесс приобретения ранее индифферентным звуковым стимулом определенного сигнального значения при выработке условного рефлекса. Специальные исследования показали, что преобразование ранее неэффективного звукового тона в мотивационпо окрашенный сигнал оборонительной реакции животного сопровождается возникновением избирательного реагирования элементов фронтальной коры именно на данную частоту заполнения тонального сигнала. Однако по мере стабилизации и закрепления условного рефлекса такая избирательность выражалась слабее и наконец исчезала, что свидетельствует о динамической роли фронтального пеокортекса в формировании условно-рефлекторной сепсомоторпой интеграции.
Описанные выше закономерности избирательного реагирования иа сигнальио значимые агенты среды могут служить физиологическими предпосылками для трактовки механизмов сенсорного внимания (Н. Ф. Суворов, О. П. Таиров, 1986).
В самом общем смысле внимание рассматривается как один из механизмов устранения избыточности сенсорных сообщений, который участвует как в избирательном регулировании сенсорных сообщений, так и в фильтрации информации, извлекаемой из систем кратковременной и долговременной памяти. Принято думать, что процессы фильтрации осуществляются уже в специфических образованиях сенсорных систем. Между тем в ряде работ мнение об уменьшении избыточности информации основано на данных об уменьшении плотности потока импульсации при переходе от нижележащих уровней сенсорных систем к вышележащим и на постулировании использования в их деятелыюсти двоичного кода. Этот вопрос, однако, остается нерешенным с нейрофизиологических позиций, и проблемой «внимания» преимущественно владеют психологи и психофизиологи.
Сам А. А. Ухтомский (1923), сформулировав принцип доминанты как физиологическую основу акта внимания, писал, что из множества действующих рецепций доминанта вылавливает группу рецепций, которая для нее в особенности биологически значима.
Определение вектора поведения (по принципу доминанты) основано иа взаимодействии биологически значимых экстероцеитивпых (окружающая среда) и иптероцептивиых (мотивационпых) воздействий, основным субстратом установления которых являются таламокортикальиые ассоциативные системы мозга.
Следовательно, целостная сенсорная функция мозга обеспечивается содружественной деятельностью сенсорных и ассоциативных систем и направлена па организацию адаптивных движений и действий. Последнее лежит в основе согласования движения с чувствованием (И. М. Сеченов, 1863).