

Филогенетические реликты
К изучению скоростей эволюции групп прямое отношение имеет проблема так называемых филогенетических реликтов (персистентных форм, или «живых ископаемых») — форм, сохраняющих неизменными основные особенности строения на протяжении огромных промежутков времени.
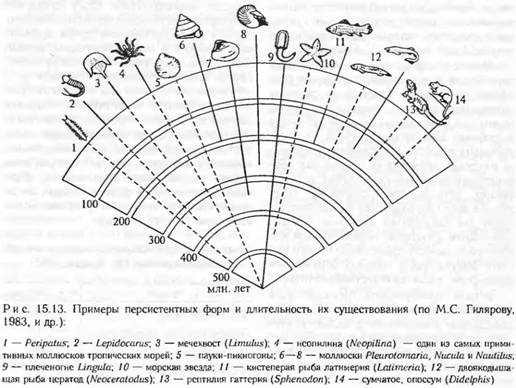
Морское плеченогое лингула (Lingula) встречается в неизменном виде, начиная с ордовика, т. е. существует на Земле как вид более 500 млн лет! Моллюск Neopilina, обитающий ныне в Тихом океане, очень похож на виды того же рода, обитавшие в нижнем девоне, т. е. не менее 400 млн лет назад. Мечехвост (Limulus) — обычный обитатель тропических прибрежных вод Юго-Восточной Азии — в почти неизменной форме существует с силура (около 400 млн лет). Знаменитая кистеперая рыба ла-тимерия (Latimeria) в почти неизменном виде сохранила строение и форму представителей группы, вымершей 200—300 млн лет назад. Число примеров можно увеличить. Все они показывают, что отдельные виды могут сохраняться на протяжении сотен миллионов лет эволюции внешне малоизмененными. В чем причина такого удивительного постоянства?
Во всех случаях оказывается, что сохранение таких персистентных (неизменных) видов возможно при стабильности основных компонентов среды
Так, особенно много филогенетических реликтов в некоторых участках Мирового океана и в тропиках, где условия существования остаются стабильными на протяжении десятков миллионов лет (рис. 15.13).
Иногда процесс вымирания рассматривается как исторически неизбежный процесс «старения вида» (Ш. Депере). Согласно этой точке зрения, каждая филогенетическая ветвь проходит стадии юности, зрелости, старости. Дойдя в силу внутренней необходимости до крайней степени специализации в фазу старости, таксон якобы должен неизбежно погибнуть, не оставив потомства. Методологическая ошибка подобного рассуждения состоит в том, что филогенетические события оцениваются с позиций изменений, характерных для совершенно другого уровня организации жизни (онтогенетического), а также в том, что вымирание оказывается не результатом исторического развития, а «старения» вида. Против такой финалистской концепции свидетельствуют факты существования достаточно многочисленных персистентных форм, равно как и факты крайне быстрого вымирания некоторых видов.
Правила эволюции филогенетических групп. «Правила» эволюции групп
Сопоставления характера развития изученных ветвей древа жизни позволили установить некоторые общие черты эволюции групп. Эти эмпирические обобщения называются «правилами макроэволюции».
Правило необратимости эволюции
(Л. Долло, 1893) гласит, что эволюция — процесс необратимый и организм не может вернуться к прежнему состоянию, уже осуществленному в ряду его предков. Так, если в эволюции наземных позвоночных на каком-то этапе от примитивных амфибий возникли рептилии, то рептилии, как бы ни шла дальше эволюция, не могут вновь дать начало амфибиям. Вернувшись в просторы Мирового океана, рептилии (ихтиозавры) и млекопитающие (киты) никогда не становятся рыбами. Можно сказать, что если какая-то группа организмов в процессе эволюции вновь «возвращается» в адаптивную зону существования ее предков, то приспособление к этой зоне у «вернувшейся» группы будет неизбежно иным.
Правило необратимости эволюции в наше время получило существенное уточнение. Успехи генетики позволяют говорить о возможности повторного возникновения признаков на основе обратных мутаций.
Но признание обратимости отдельных признаков (П
П. Сушкин и др.) в филогенезе — отнюдь не признание обратимости эволюционного процесса в целом. Обратная мутация по признаку может привести к повторному возникновению данного аллеля, но не генотипа в целом; к вторичному появлению данного признака, но не фенотипа в целом. Статистически вероятно повторное возникновение мутаций, но статистически невероятно повторное возникновение генных комплексов и целых фенотипов (Н.Н. Воронцов).
Правило прогрессирующей специализации (Ш. Депере, 1876) гласит, что группа, вступившая на путь специализации, как правило, в дальнейшем развитии будет идти по пути все более глубокой специализации.
Если в процессе эволюции одна из групп позвоночных, скажем ветвь рептилий, приобрела адаптации к полету, то на последующем этапе эволюции это направление адаптации сохраняется и усиливается (например, птеродактили в свое время все более приспосабливались к жизни в воздухе). Поскольку организм определенного строения не может жить в любой среде, в выборе адаптивной зоны или ее части группа ограничена особенностями строения. Если эти особенности несут черты специализации, то организм обычно
«выбирает» (точнее, в результате борьбы за существование попадает во все более частную среду), где его специализированные приспособления могут обеспечить успешное выживание и оставление потомства. Но обычно это ведет лишь к дальнейшей специализации.
Частный случай этого общего правила прогрессивной специализации — увеличение размеров тела особей в процессе эволюции позвоночных животных. Увеличение размеров тела, с одной стороны, связано с более экономным обменом веществ (уменьшение величины относительной поверхности тела) и должно рассматриваться как частный случай специализации. С другой стороны, дает хищнику преимущества в нападении, а жертве — преимущества в защите. Связь организмов в цепях питания неизбежно вызывает увеличение размеров тела во многих группах. У представителей других групп происходит уменьшение размеров тела. Например, при переходе к подземному образу жизни и обитанию в закрытых норах многие грызуны вторично стали более мелкими. Интересно, что сопряженной эволюции подверглась ласка (Mustela nivalis) — один из наиболее облигатных потребителей мышевидных грызунов в средней полосе. Ласка приобрела размеры тела, позволяющие преследовать мелких грызунов в норах
На этом примере видно, что выделяемые эмпирические эволюционные правила имеют относительное значение. Характер эволюции зависит в конечном счете от конкретных связей группы с эле-ментами биотической и абиотической среды (всегда при постоянном контроле отбора, идущем на уровне микроэволюционных взаимодействий внутри популяций и биогеоценозов).
Правило происхождения от неспециализированных предков (Э. Коп, 1896)гласит, что обычно новые крупные группы берут начало не от специализированных представителей предковых групп, а от сравнительно неспециализированных. Млекопитающие возникли не от высокоспециализированных форм рептилий,а от неспециализированных. Голосеменные растения возникли от неспециализированных палеозойских папоротникообразных. Ныне процветающая группа цветковых растений возникла не от специализированных голосеменных, а от неспециализированных предков, занимающих промежуточное положение между семенными папоротниками и беннетитовыми (см. гл. 6).
Причина происхождения новых групп от неспециализированных предков в том, что отсутствие специализации определяет возможность возникновения новых приспособлений принципиально иного характера
Трудно ожидать появления среди гельминтов каких-то принципиально новых форм. У таких форм скорее могут возникнуть эволюционные изменения, направленные к лучшему прикреплению внутри организма хозяина, лучшему использованию питательных веществ, более эффективному способу размножения и т. п. Напротив, у такой группы, как практически всеядные средних размеров хищные, живущие в разнообразных условиях, большие потенциальные возможности развития в самых разных направлениях.
Высокая потентность неспециализированных групп в эволюции определяется борьбой за существование, которую они вынуждены вести. Но в историческом плане именно эти жесткие требования среды и приводят потомков неспециализированных групп на путь арогенеза, вызывая к жизни приспособления, которые потом оказываются перспективными для начала нового широкого аллогенеза (см. гл. 15).
Правило происхождения от неспециализированных предков оказывается, однако, далеко не всеобщим. Редко бывает так, чтобы специализация затронула в равной степени все без исключения органы и функции организма (вида)
Эти остающиеся достаточно лабильными органы и системы могут открыть группе дорогу в другую адаптивную зону. Переход к птицам был осуществлен в группе рептилий, уже способных к лазанию и прыганию по ветвям деревьев, т. е. в специализированной с точки зрения развития приспособлений к передвижению группе. Переход к наземному образу жизни среди позвоночных был осуществлен формами, способными, по-видимому, передвигаться по твердому субстрату, заглатывать и использовать для дыхания кислород воздуха, т. е. опять-таки формами, крайне специализированными в этом отношении по сравнению с большинством обитавших в водоемах видов рыб. Оказывается, признаки, специализированные в одной среде, могут быть эрогенными в иной (М.С. Гиляров). Ясно, что как неспециализированные, так и специализированные формы могут при определенных условиях стать родоначальниками новых, прогрессивных ветвей древа жизни.
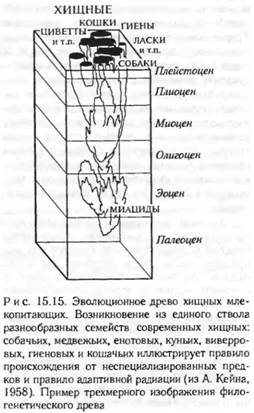
Правило адаптивной радиации (Г.Ф. Осборн, 1902) гласит, что филогенез любой группы сопровождается разделением группы на ряд отдельных филогенетических стволов, которые расходятся в разных адаптивных ходит за пределы узких условий
Другие дают возможность выхода группы в новую адаптивную зону и непременно ведут к быстрому эволюционному развитию групп в новом направлении. По существу, это правило не что иное, как принцип дивергенции, подробно описанный Ч. Дарвином (1859) при обосновании гипотезы естественного отбора. Дарвин говорил о внутривидовой приспособительной дивергенции к различной пище, несколько различным условиям существования и т. п. и рассматривал ее как обязательный этап образования новых видов. В дальнейшем принцип дивергенции был положен в основу представлений эволюционной морфологии об идиоадаптациях (А.Н. Северцов) и соответственно о развитии группы по пути аллогенеза, а в эволюционной палеонтологии — в основу представлений об «иррадиации» (В.О. Ковалевский, 1875) в процессе становления каждого крупного филума (рис. 15.15, см. рис. 6.4) и классов в систематике (Дж. Хаксли).
Правило чередования главных направлений эволюции. Арогенная эволюция чередуется с периодами аллогенной эволюции во всех группах. И.И. Шмальгаузен (1939) сформулировал это правило как чередование фаз адаптациоморфоза.
Эволюция представляет непрерывный процесс возникновения и развития новых и новых адаптации — адаптациогенез.
Обычно одна из форм получает какую-то новую адаптацию (или комплекс адаптации), оказывающуюся основой для следующего быстрого периода филогенетических новообразований (рис. 15.16).
Правило усиления интеграции биологических систем (И.И. Шмальгаузен, 1961) можно сформулировать так: биологические системы в процессе эволюции становятся все более интегрированными, со все более развитыми регуляторными механизмами, обеспечивающими такую интеграцию.
Сейчас в общих чертах известны основные направления такой интеграции, идущей на уровне популяции и биогеоценозов. На уровне популяции это означает поддержание определенного уровня гетерозиготности, которая является основой интеграции всего популяционного генофонда в сложную, лабильную и одновременно устойчивую генетическую систему, способную к саморегуляции (численности, структуры), ге-нетико-экологическому гомеостазису.
Межпопуляционные взаимоотношения интегрируют отдельные популяции и их группы в вид как систему (см. гл. 12, 13). Интеграция в системе биогеоценоза определяет образование разными видами многих взаимодополняющих друг друга экологических ниш и слаженной работой биогеоценоза как элементарной биохорологической структуры в биосфере нашей планеты (с упорядоченными потоками вещества, энергии, информации). Высший уровень интеграции выражается в эволюции самих механизмов эволюции — проблема, которая начинает вырисовываться в современном эволюционном учении.
Несомненно, возможно выделение и других кроме шести выделенных нами выше «правил». Тут открыто широкое поле деятельности для исследователя-эволюциониста.