

Дополнительная моторная область
Дополнительная моторная область имеет, однако, иную топографическую организацию для контроля двигательной функции. Эта область лежит главным образом в продольной щели большого мозга, но занимает также несколько сантиметров верхней поверхности лобной коры. Сокращения, вызываемые стимуляцией этой области, — часто не односторонние, а двусторонние. Например, стимуляция ведет к одновременному сжиманию кистей обеих рук; такие движения, вероятно, являются рудиментарной функцией кистей, необходимой для лазания.
В целом эта область функционирует совместно с премоторной корой для обеспечения общей двигательной активности, необходимой для поддержания равновесия, фиксации определенных сегментов тела, позных движений головы и глаз и других, лежащих в основе более тонкого двигательного контроля рук и кистей премоторной областью и первичной моторной корой.
Нейрохирурги обнаружили у человека несколько высокоспециализированных моторных областей коры большого мозга, которые контролируют специфические двигательные функции. Локализация этих регионов была установлена либо путем электрической стимуляции, либо на основании данных о выпадении моторных функций при деструктивных поражениях определенных корковых областей. Некоторые из наиболее важных регионов следующие.
Область Брока и речь. Премоторная область, обозначенная «формирование слов» и лежащая непосредственно впереди первичной моторной коры и прямо над сильвиевой щелью - это область Брока. Ее поражение не лишает человека возможности издавать некоординированные звуки, восклицания или изредка такие простые слова, как «нет» или «да», но делает невозможным произнесение целых слов. Корковая область, тесно связанная с областью Брока, обеспечивает также соответствующую дыхательную функцию, благодаря которой одновременно с движениями рта и языка во время речи происходит дыхательная активация голосовых связок.
Поле регуляции для произвольных движений глаз. В премоторной области, непосредственно выше области Брока, есть участок для регуляции произвольных движений глаз. При поражении этой области человек не может произвольно направлять взор на различные объекты. Вместо этого проявляется тенденция к непроизвольной фиксации глаз на определенных объектах — эффект, контролируемый сигналами из затылочной зрительной коры. Эта лобная область также контролирует движения век, например мигание.
Область вращения головы. Электрическая стимуляция моторной ассоциативной области, расположенной несколько выше, вызывает поворот головы. Эта область тесно связана с областью движения глаз; она поворачивает голову в направлении определенных объектов.
Область для тонкой работы руками (моторный центр праксиса). В премоторной коре непосредственно перед первичной моторной корой есть область, которую нейрохирурги определили как важную для ручных навыков (область кистей и пальцев рук). При разрушении этой области опухолью или по другим причинам движения кистей рук становятся некоординированными и нецеленаправленными. Такое состояние называют моторной апраксией.
Двигательные пути, идущие от головного мозга к спинному, делятся на две системы: пирамидную и экстрапирамидную.
Примерно 30% волокон кортикоспинального тракта исходят из первичной моторной коры, 30% — от премоторной и дополнительной моторной коры, а 40% — от соматосенсорных областей, расположенных позади центральной борозды.
После выхода из коры тракт проходит через заднюю ножку внутренней капсулы (между хвостатым ядром и скорлупой базальных ганглиев) и спускается вниз через ствол мозга, формируя пирамиды продолговатого мозга. Основная часть волокон пирамидного тракта в нижней части продолговатого мозга перекрещивается, переходя на противоположную сторону и спускаясь в составе латеральных кортикоспинальных трактов спинного мозга. В основном эти волокна формируют синапсы на вставочных нейронах промежуточных областей серого вещества спинного мозга. Часть волокон заканчиваются на сенсорных релейных (переключающих) нейронах в задних рогах, и очень небольшое число волокон синаптически связаны непосредственно с передними мотонейронами, которые вызывают сокращение мышц.
Пирамидный тракт: начинаясь в моторной и сенсомотрной зонах коры больших полушарий, большая часть волокон направляется прямо к эфферентным нейронам в передних рогах спинного мозга.
Экстрапирамидный тракт идет к передним рогам спинного мозга, передает им эфферентную импульсацию, обработанную в комплексе подкорковых структур (базальных ганглиях, таламусе, мозжечке).
Небольшая часть кортикоспинальных волокон не переходит на противоположную сторону в продолговатом мозге, а проходит по той же стороне (ипсилатерально) спинного мозга в вентральных кортикоспинальных трактах. Многие из волокон, если не большинство, в итоге тоже переходят на другую сторону спинного мозга на уровне шейных или верхних грудных сегментов. Эти волокна, возможно, связаны с регуляцией билатеральных позных движений, управляемых дополнительной моторной корой.
Наиболее впечатляющей является популяция крупных мякотных волокон пирамидного тракта со средним диаметром 16 мкм. Эти волокна исходят от гигантских пирамидных клеток, называемых клетками Беца, которые обнаруживаются только в первичной моторной коре. Диаметр клеток Беца — примерно 60 мкм, а их волокна проводят импульсы к спинному мозгу со скоростью около 70 м/сек (наибольшая скорость проведения сигналов от головного к спинному мозгу).
В каждом кортикоспинальном тракте примерно 34000 волокон от клеток Беца. Общее число волокон в каждом тракте — более 1 млн, и эти крупные волокна составляют лишь 3% общего числа, а остальные 97% представлены в основном волокнами диметром менее 4 мкм, которые проводят фоновые тонические сигналы в двигательные области спинного мозга.
Другие пути волокон от моторной коры. Моторная кора дает начало большому числу дополнительных, главным образом тонких, волокон, которые идут к глубоким регионам большого мозга и мозгового ствола, включая следующие.
1. Аксоны гигантских клеток Беца посылают короткие коллатерали к самой коре. Считают, что эти коллатерали тормозят прилежащие регионы коры, когда разряжаются клетки Беца, «обостряя» таким образом границы возбудительного сигнала.
2. Большое число волокон проходит от моторной коры в хвостатое ядро и скорлупу. Отсюда дополнительные пути идут в ствол мозга и спинной мозг, что обсуждается в следующей главе, главным образом для регуляции сокращений постуральных мышц тела. 3. Умеренное количество двигательных волокон идут к красному ядру среднего мозга. От них волокна проходят вниз к спинному мозгу по руброспиналъному тракту.
4. Умеренное число двигательных волокон заходят в ретикулярную формацию и вестибулярные ядра ствола мозга; отсюда сигналы идут к спинному мозгу по ретикуломозжечковым и вестибуломозжечковым трактам. 5. Огромное количество моторных волокон синаптически прерываются в ядрах моста, которые дают начало мостомозжечковым волокнам, несущим сигналы в полушария мозжечка.
6. Коллатерали также заканчиваются в ядрах нижней оливы и отсюда вторичные оливомозжечковые волокна передают сигналы к множественным областям мозжечка. Таким образом, каждый раз, когда сигнал передается вниз к спинному мозгу для вызова двигательной активности, базальные ганглии, ствол мозга и мозжечок получают мощные сигналы от кортикоспинальной системы.
|
|
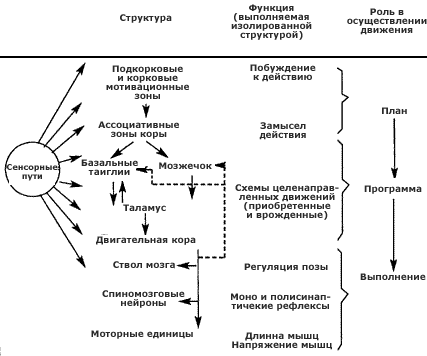
Общий план организации двигательной системы (по Дж. Дуделу с соавт., 1985).
Важнейшие двигательные структуры и их основные взаимосвязи указаны в левом столбце. В среднем столбце перечислены самые главные и твердо установленные функции, обнаруженные при раздельном изучении каждой из этих структур. В правом столбце указано, каким образом эти функции связаны с возникновением и выполнением движения.