

Глава VI
Нейрофизиологическое исследование эмоциональных процессов у детей
Одной из важнейших проблем, где встречаются научные интересы психологии и нейрофизиологии, является проблема формирования у человека социальных эмоций.
Приступая к изучению нейрофизиологических механизмов социальных эмоций у детей, следует в первую очередь понять, каков генез этих процессов, какую функцию выполняют эмоции в регуляции поведения ребенка, как связаны эти сложные образования с развитием его когнитивной сферы — речи и мышления.
На важную роль эмоций в регуляции мыслительных процессов указывали С. Л. Рубинштейн [1946], А. Н. Леонтьев [1977] и другие. Подлинной конкретной единицей психического, считал С. Л. Рубинштейн, является целостный акт отражения объекта субъектом. Это отражение, сложное по составу, всегда в той или иной мере включает единство двух противоположных компонентов — знания и отношения, интеллектуального и аффективного, из которых то один, то другой выступают в качестве преобладающего. Мышление как деятельность, указывал А. Н. Леонтьев [1977], имеет и аффективную регуляцию, непосредственно выражающую ее пристрастность.
Эти положения, разработанные советскими психологами, являются основополагающими; они ориентируют исследователей, дают ключ к поиску путей изучения нейрофизиологических механизмов социальных эмоций у детей.
С какими же физиологическими процессами может быть связано развитие таких сложных эмоций, как социальные чувства детей, опосредствованные деятельностью мышления, какие корковые системы могут быть ответственны за регуляцию эмоционального поведения ребенка, какова роль правого и левого полушарий в развитии эмоций — эти и другие вопросы являются почти не изученными в науке. Особый интерес среди этих вопросов представляет изучение роли коры больших полушарий, являющейся ведущим интегрирующим звеном всех мыслительных процессов.
Многочисленными клиническими и нейропсихологи-ческими исследованиями установлено, что с функциями коры больших полушарий головного мозга связаны наи-более сложные и тонко организованные формы эмоционального поведения человека. При корковых поражениях развиваются патологические виды поведения — растормаживаются врожденные эмоции, влечения. Больные становятся прожорливыми, эротичными, у них появляются тяжелые психические расстройства от аффектов чрезвычайной силы до состояния глубокой апатии,
своеобразного эмоционального паралича. Особое зна- чение имеют лобные отделы неокортекса в регуляции специфически человеческих форм эмоционального пове- дения.
П. К. Анохин [1968] указывал на то, что лобные отделы мозга являются структурой, где первичный мотивационный признак стимула может быть превращен в организующий, ведущий к формированию функциональ- ной системы. В экспериментальных исследованиях на животных показано, что лобная кора непосредственно связана с организацией целенаправленного поведения, с проявлениями мотивационных процессов, с перераста-нием одной из наличных потребностей в доминирующую мотивацию и удержанием этой мотивации в течение определенного периода времени. Лобные доли, таким образом, играют значительную роль в формировании и стабилизации доминирующей мотивации [Ю. В. Уры-ваев, 1979].
Разрабатывая информационную теорию эмоций, П. В. Симонов [1979, 1981] указывает, что передние отделы лобной коры связаны с ориентацией поведения животных на сигналы с высокой вероятностью их подкрепления. После удаления этих отделов значение сигналов как бы выравнивается, и животное начинает одинаково реагировать на сигналы с высоко- и маловероятным подкреплением. Лобные области являются высшим корковым уровнем регуляции эмоционально-приспособительного поведения животных [А. Я. Мехедова, 1972]. В работе С. П. Шклярук с соавторами [1979] показано, что поляризация лобной коры обезьян вызывает у них существенные перестройки эмоционального состояния. Развитие лобных долей обезьян, как считает Ш. Л. Джалагония [1969], биологически обусловлено выполнением на более высоком уровне функций, лежащих в основе сложных и эмоционально насыщенных форм поведения приматов. У обезьян нарушение цингу-лярной и дорзолатеральной префронтальной коры со- провождается укорочением реакции избегания без изменения агрессивности [К. Pribram, J. Fulton, 1954] и повышением порога фрустрационных реакций [J. Stamm, 1964]. Последний факт позволил автору сделать вывод о вовлечении этих областей новой коры в сложный нейрональный контроль эмоционального поведения. В целом экспериментальные исследования, выполненные на животных, показали, что эмоциональные процессы
тесно связаны с функциями коры больших полушарий головного мозга, и в первую очередь с включением передних ассоциативных лобных долей в управление этими процессами.
Клиническими наблюдениями установлено, что лобные доли мозга имеют особое значение в детерминировании специфических форм эмоционального поведения человека. При поражении этих долей наибольший ущерб терпят процессы эмоциональных переживаний, связанных с познавательными функциями, с социальным поведением, и вместе с тем начинают доминировать пассивные, биологически неадекватные эмоциональные проявления в виде тупой эйфории, благодушия, беспечности, потери чувства такта, грубой сексуальной расторможен-ности или, напротив, безразличия к окружающим, вялости и апатии [А. Р. Лурия, 1966; Т. А. Доброхотова,, 1974 и др.].
Каким образом эмоциональные процессы у человека развиваются и совершенствуются в онтогенезе в связи? с развитием речевого мышления? Мы предприняли первую попытку в изучении этого вопроса, важного как для» разработки проблем теории эмоций, так и практики воспитания и развития личности ребенка.
Была поставлена задача изучить у детей в возрасте 3 — 6 лет нейрофизиологические механизмы социальных эмоций, опосредованных речевым мышлением, и выявить роль передних и задних ассоциативных зон коры больших полушарий в их регуляции.
1. Ассоциативные отделы неокортекса и их роль в оценке значения и смысла речевых сигналов
При изучении нейрофизиологических механизмов социальных эмоций у детей, связанных с процессами речевого мышления, с формированием сознания собственного «Я», необходимого, чтобы «примерить» к нему чужие беды, радости, переживания, мы вплотную сталкиваемся с проблемой смыслов. Еще Л. С. Выготский [1982] различал понятия «значение» — объективное отражение обобщенных связей и отношений действительности — и. «смысл» — выбор из всех возможных систем значений, только тех сторон, которые соответствуют потребности, субъекта, представляют для него специальный интерес. В этом проявляется одна из важных особенностей пси-
хических процессов — их избирательность, целенаправ- ленность.
Нейрофизиологические исследования показали, что выполнение одного и того же действия или восприятие одной и той же информации осуществляются мозгом по- разному, в зависимости от того, какое значение (смысл) имеют эти действия или информация для ребенка.
Рассмотрим конкретные примеры. Корреляционный анализ ЭЭГ, записанной у детей в возрасте 5—6 лет при восприятии рассказа «На льдине», позволил установить, что функциональная организация межцентральных взаимодействий проекционных и ассоциативных корковых зон определяется не только объективным значением рече- вых сигналов, поступающих по сенсорным каналам, но и их смыс- лом, эмоциональной значимостью, отношением ребенка к конкрет- ной речевой деятельности. Так, оказалось, что наиболее существен- ные сдвиги в общей организации межцентральных взаимодействий отмечались на слово медведи только тогда, когда оно приобретало для детей эмоциональную значимость (медведи могут напасть на детей, которых унесло на льдине в море), когда после осмыслива- лия ситуации возникало эмоциональное отношение к героям рас- сказа. При этом почти в 1,5 раза повышалась степень межцентраль- ных взаимодействий (рис. 1), усиливались взаимосвязи височных зон, т. е. повышалась функциональная активность специфических сенсорных входов, наиболее заинтересованных в восприятии речи. Более чем в 2 раза повышалась функциональная активность задних ассоциативных нижнетеменных зон и более чем в 7 раз возрастал уровень активности передних ассоциативных лобных зон левого по- лущария, т. е. наблюдалась межполушарная асимметрия.
Приведенные факты свидетельствуют, по-видимому,
о том, что лобные зоны активно включались в процесс
межсенсорной интеграции и обеспечивали не только
оценку значения слова, но и понимание смысла речево-
го сообщения. При этом повышались уровень и степень
межцентральных взаимодействий как в правом, так и
левом полушарии, причем в левом более значительно.
Кроме того, усиливались процессы взаимодействия меж-
ду полушариями за счет увеличения «асимметричных»
межполушарных связей, особенно нижнетеменной зоны
обоих полушарий.
Важным в теоретическом плане оказался факт уси-
ления активности исследованных структур мозга и пе-
рераспределения межцентральных взаимодействий не на
само слово, а еще до его звучания при повторении рас-
сказа, т. е. когда ребенок уже знал смысл всего речево-
го сообщения. Ребенок эмоционально предвосхищал по-
следующие события, и его мотивационно-смысловая
ориентировка осуществлялась в плане воображения, а
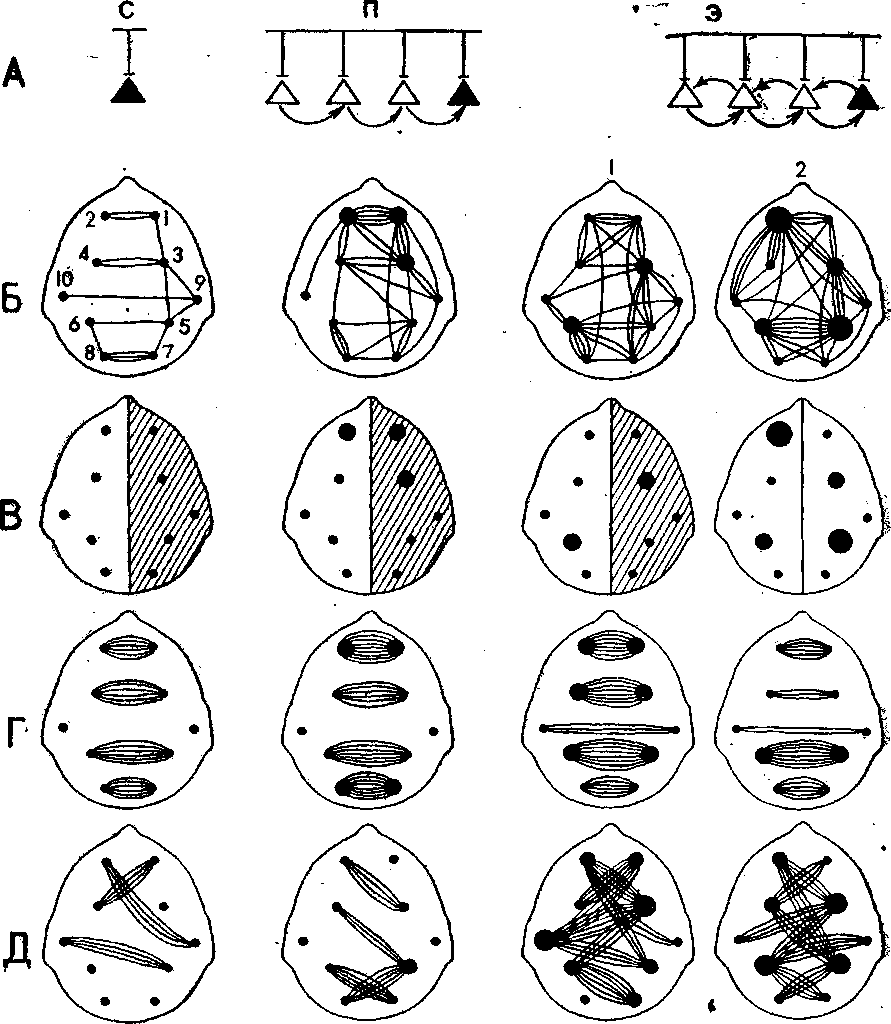
Рис. 1. Межцентральные взаимодействия в коре головного мозга у детей при восприятии слова медведи, приобретающего разный смысл. А — схема усложнения значений слова медведи в эксперименте; Б — суперпозиция высокозначимых (г≥0,7) корреляций биопотенциалов мозга (ЭЭГ) по группе детей; В — доминирующее полушарие по уровню пространственной синхронизации биопотенциалов мозга (показано штриховкой), активность отдельных корковых зон пропорциональна диаметру кружков; Г — суперпозиция значимых (г ≥0,4) межполушарных корреляций симметричных центров по группе детей; Д — то же для асимметричных центров; С — во время восприятия слова медведи вне текста рассказа, П — того же слова в познавательной части рассказа, Э — того же слова в эмоциональной части рассказа (1 — перед восприятием слова, 2 — во время восприятия). На корреляционных схемах цифрами обозначены области отведения ЭЭГ; 1 и 2 — лобные, 3 и 4 — моторные, 5 и 6 — нижнетеменные, 7 и 8 — затылочные», 9 и 10 — височные.
не в плане непосредственного восприятия. Эмоции на- чинали по времени опережать действие раздражителя, момент восприятия ключевого слова сообщения. Ребе- нок, становясь на позицию героев, начинал им сопере- живать. В этот момент эмоционального предвосхищения ситуации наблюдалось возрастание степени меж- центральных взаимодействий, особенно это было харак- терно для нижнетеменных областей, участвующих, по- видимому, в репродукции эмоционального образа. Зна- чительно повышался уровень межполушарных взаимо-действий, особенно за счет увеличения «асимметричных» связей левой височной области, специфически ответст- венной за реализацию речевой деятельности, за хране- ние и извлечение из памяти слуховой информации.
Таким образом, «включение» процессов речевого мышления позволяло ребенку осмыслить ситуацию в це- лом и соответственно по-разному реагировать на звуча- ние одного и того же слова, которое имело для него разный смысл и, естественно, разную эмоциональную значимость.
Тесную связь высших эмоций с речевым мышлением
демонстрирует и пример, когда дети 3 лет на ощупь
опознавали предметы, имеющие разный смысл
[Т. P. Khrisman, V. D. Eremeeva, 1984]. В первый мо-
мент при восприятий предмета (пушистого, мягкого) на
ощупь, т. е. при формировании его первичного перцеп-
тивного образа, наиболее функционально активной бы-
ла задняя ассоциативная нижнетеменная область, где
выявлялись фокусы взаимосвязанной активности и уси-
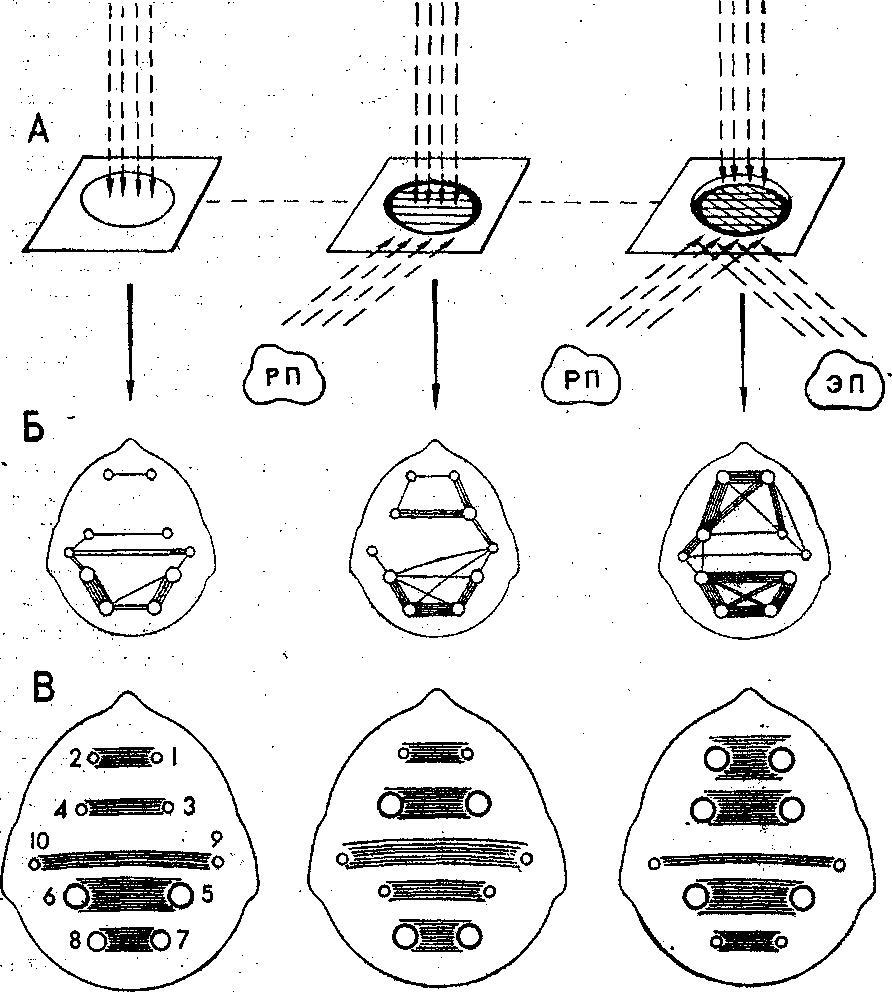
ливались межполушарные взаимодействия (рис. 2). От-
мечалось доминирование теменно-затылочных связей.
После того как этот предмет был вербализован («Это
лапка»), т. е. когда ему придали конкретное значение,
что, вероятно, должно было способствовать формирова-
нию более целостных перцептивных образов и их более
активному переносу в план воображения, можно было
ожидать и включения более высокого уровня интегра-
ции поступающей сенсорной информации. Подтвержде-
ние этому было обнаружено и в характере изменений в
картине межцентральных взаимодействий. Так, в этот
момент усилились межполушарные моторно-моторные
взаимодействия (в 67% случаев) и повысилась степень
межцентральных связей моторной зоны правого полу-
шария. Иное значение, а главное, иной смысл приобрел
этот предмет, когда ребенку после просмотра сказки
! II II!
5—1655
«Красная Шапочка» сказали: «Это лапа Волка», т.е. когда смысл данного предмета, обозначенный через слово, обусловливал выраженную эмоциональную peaк- цию ребенка при восприятии этого предмета. Только в данном случае отмечалось повышение функциональной активности передних ассоциативных лобных зон. При этом усиливались межцентральные отношения симмет- ричных центров лобных и нижнетеменных зон, повы- шалея уровень лобно-моторных связей в левом полуша- рии, и в этих зонах выявился фокус наибольшей взаи- мосвязанной активности.
Итак, изменению посредством слова смысла воспри- нимаемой ребенком по разным сенсорным каналам ин- формации, что обусловливало те или иные изменения его эмоционального отношения к ней, соответствовали и перестройки в функциональных межцентральных взаи- модействиях, особенно ассоциативных отделов лобных и нижнетеменных зон.
Еще более доказательно тесная зависимость эмоций ребенка от смысла деятельности была показана при ее оценках, разных по знаку и отнесенных к ее разным фрагментам. При этом предполагалось, что ярко выра- женными эмоции в ответ на оценку будут в том случае, когда будет оцениваться самый важный, центральный фрагмент деятельности, в котором был заключен для ребенка ее смысл. Деятельность детей «сенсорной» группы, только слушавших рассказ, оценивалась слова- ми «плохо — хорошо слушал», а «моторной», слушавших и в игровой ситуации активно выполнявших роль радиста, — «плохо — хорошо слушал» и «плохо — хорошо выполнял». Следует также отметить большую конкретность и четкую отнесенность к определенному фрагменту деятельности оценок «плохо — хорошо выполнял».
Анализ особенностей поведения детей, частоты появ- ления и мощности КХР показал, что для детей «мотор- ной» группы оценки «плохо — хорошо выполнял» оказались гораздо более эмоционально значимы, чем «плохо — хорошо слушал». Такая дифференцированность в выраженности эмоциональных переживаний у этих детей свидетельствовала, что именно активные действия в роли радиста приобретали для них ведущий смысл. Его выделение было обусловлено как сюжетной линией рассказа, так и специальными инструкциями экспериментатора и облегчалось наличием внешнего плана деятельности.
«ПЛОХО...»
«ХОРОШО...» «ПЛОХО...» «ХОРОШО...»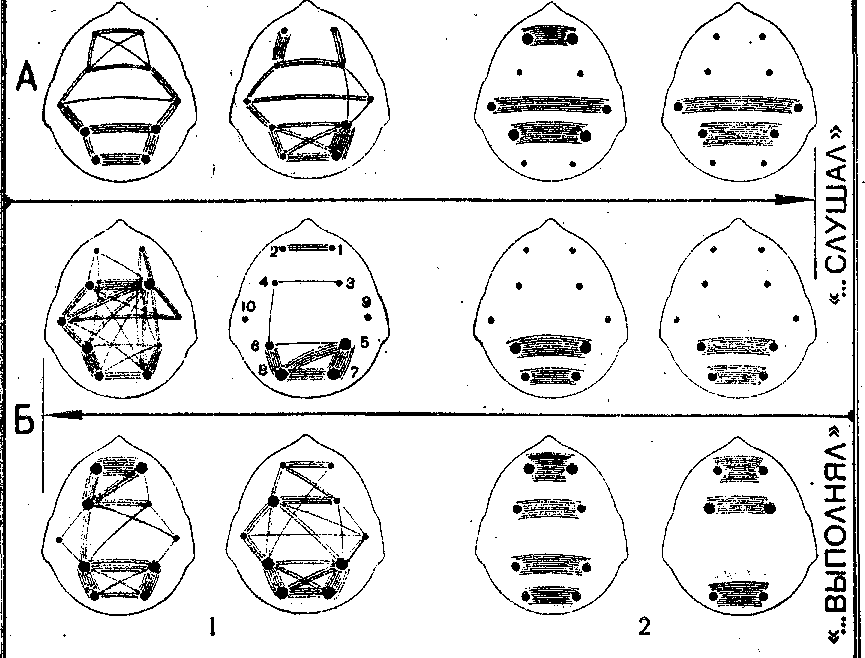
А — группа детей, которые слушали рассказ;. Б — группа детей, кото- рые не только слушали рассказ, но и выполняли в нем игровую роль радиста. Оценки «плохо — хорошо слушал» давались детям обеих групп, а «плохо — хорошо выполнял» — только детям, игравшим в рассказе роль радиста. 1 — суперпозиции высокозначимых (r≥ 0,7) корреляций биопотенциалов мозга (ЭЭГ) по группам детей, 2 — суперпозиции значимых (г≥0,4) межполушарных корреляций симметричных центров по группам детей. На корреляционных схемах цифрами от 1 до 10 обозначены отведения ЭЭГ, как на рис. 1.
Именно в моменты восприятия оценок «плохо — хорошо выполнял» по сравнению с «плохо — хорошо слушал» у детей «моторной» группы наблюдалась наибольшая активность лобных и моторных центров коры (рис. 3). При этом повышались общий уровень и степень синхронизации потенциалов мозга, особенно симметричных центров лобных и моторных зон. Примерно в 2 раза повышался уровень межполушарных взаимодействий лобных центров (с 50 и 53,3% случаев на оценки «плохо— хорошо слушал» до, соответственно, 86,7 и 100% на оценки «плохо — хорошо выполнял»). В лобной (на отрицательную оценку) и моторной зонах слева появля-
5*
лись фокусы наибольшей взаймосвязанной активности и выявлялась левосторонняя асимметрия.
У детей «сенсорной» группы в момент восприятия оценок «плохо — хорошо слушал» наблюдалась иная специфика функциональной активности корковых цент- ров мозга. При этом отмечался наиболее высокий уро- вень связей симметричных центров лобных зон в момент отрицательной оценки и его снижение при положитель- ной (с 71,4% до 43,8%). Здесь следует подчеркнуть, что этим детям была свойственна и более выразительная экспрессия на отрицательную оценку, чем на положи- тельную. Характерной же чертой в картине межцент- ральных взаимодействий при восприятии оценок у де- тей «сенсорной» группы была относительно высокая и стабильная функциональная активность нижнетеменных и височных зон мозга.
Особо интересным представляется тот факт, что в моменты восприятия оценок избирательно усиливалась функциональная активность тех центров мозга, которые были специфически «заинтересованы» в реализации оцениваемой деятельности — височных и моторных центров соответственно у детей «сенсорной» и «моторной» групп. Более того, можно было отметить принципиальное сходство между теми паттернами активности мозга, которые обнаруживались в моменты реальной деятельности, и теми, которые появлялись при оценках этих моментов деятельности, когда она была уже завершена. У детей «моторной» группы данное сходство было выражено явно, что, видимо, обусловливалось более конкретным и направленным характером оценок «плохо — хорошо выполнял». В целом можно отметить важность полученных фактов в плане объективной диагностики ключевых, смысловых фрагментов деятельности ребенка, в плане прогноза корректирующего значения оценки в его поведении.
Следует подчеркнуть, что у детей регулирующая функция эмоциональных процессов, формирующихся в ответ на вербальную оценку их деятельности, имеет в своей нейрофизиологической основе функциональную активность ассоциативных, особенно лобных, зон мозга, которые, видимо, связаны с непосредственным отношением к оценке смысла любого вида деятельности.
Многочисленными клиническими, физиологическими и нейропсихологическими исследованиями установлена ведущая роль ассоциативных лобных областей в регуля-
ции сложных целенаправленных форм психической дея-тельности, в формировании стойких мотивов и целей, составлении сложных программ поведения [А. Р. Лурия; 1966, 1973, 1982]; У животных при поражении лобных долей нарушенными оказываются также более сложные формы избирательного, целенаправленного поведения [A.С. Батуев, 1973; И. М. Айвазашвили, 1974; и др.]. Появились новые электрофизиологические данные, доказывающие особую роль этих ассоциативных зон в оценке биологической значимости поступающих сигналов как у животных, так и у человека [Г. А. Куликов с соавт., 1979; S. Kojima, 1980; А. С. Батуев и соавт.,1979; А. Н. Пирогов, 1979; М. Watanade, 1981; Н. J. Leubuscher,1981].